

On the origins of Mendelian disease genes in man: the impact of gene duplication
- PMID: 21705381
- PMCID: PMC3709195
- DOI: 10.1093/molbev/msr111
On the origins of Mendelian disease genes in man: the impact of gene duplication
Erratum in
- Mol Biol Evol. 2012 Sep;29(9):2284
Abstract
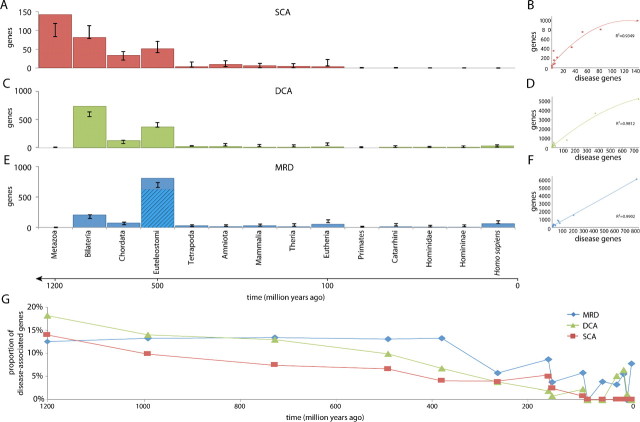
Over 3,000 human diseases are known to be linked to heritable genetic variation, mapping to over 1,700 unique genes. Dating of the evolutionary age of these disease-associated genes has suggested that they have a tendency to be ancient, specifically coming into existence with early metazoa. The approach taken by past studies, however, assumes that the age of a disease is the same as the age of its common ancestor, ignoring the fundamental contribution of duplication events in the evolution of new genes and function. Here, we date both the common ancestor and the duplication history of known human disease-associated genes. We find that the majority of disease genes (80%) are genes that have been duplicated in their evolutionary history. Periods for which there are more disease-associated genes, for example, at the origins of bony vertebrates, are explained by the emergence of more genes at that time, and the majority of these are duplicates inferred to have arisen by whole-genome duplication. These relationships are similar for different disease types and the disease-associated gene's cellular function. This indicates that the emergence of duplication-associated diseases has been ongoing and approximately constant (relative to the retention of duplicate genes) throughout the evolution of life. This continued until approximately 390 Ma from which time relatively fewer novel genes came into existence on the human lineage, let alone disease genes. For single-copy genes associated with disease, we find that the numbers of disease genes decreases with recency. For the majority of duplicates, the disease-associated mutation is associated with just one of the duplicate copies. A universal explanation for heritable disease is, thus, that it is merely a by-product of the evolutionary process; the evolution of new genes (de novo or by duplication) results in the potential for new diseases to emerge.
Figures



References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
