

Quantification of inbreeding due to distant ancestors and its detection using dense single nucleotide polymorphism data
- PMID: 21705750
- PMCID: PMC3176119
- DOI: 10.1534/genetics.111.130922
Quantification of inbreeding due to distant ancestors and its detection using dense single nucleotide polymorphism data
Erratum in
- Genetics. 2012 Jan;190(1):283
Abstract
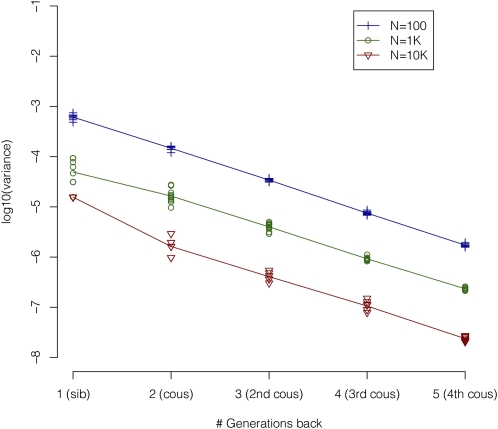
Inbreeding depression, which refers to reduced fitness among offspring of related parents, has traditionally been studied using pedigrees. In practice, pedigree information is difficult to obtain, potentially unreliable, and rarely assessed for inbreeding arising from common ancestors who lived more than a few generations ago. Recently, there has been excitement about using SNP data to estimate inbreeding (F) arising from distant common ancestors in apparently "outbred" populations. Statistical power to detect inbreeding depression using SNP data depends on the actual variation in inbreeding in a population, the accuracy of detecting that with marker data, the effect size, and the sample size. No one has yet investigated what variation in F is expected in SNP data as a function of population size, and it is unclear which estimate of F is optimal for detecting inbreeding depression. In the present study, we use theory, simulated genetic data, and real genetic data to find the optimal estimate of F, to quantify the likely variation in F in populations of various sizes, and to estimate the power to detect inbreeding depression. We find that F estimated from runs of homozygosity (Froh), which reflects shared ancestry of genetic haplotypes, retains variation in even large populations (e.g., SD=0.5% when Ne=10,000) and is likely to be the most powerful method of detecting inbreeding effects from among several alternative estimates of F. However, large samples (e.g., 12,000-65,000) will be required to detect inbreeding depression for likely effect sizes, and so studies using Froh to date have probably been underpowered.
Figures










References
-
- Abaskuliev A. A., Skoblo G. V., 1975. Inbreeding, endogamy and exogamy among relatives of schizophrenia patients. Genetika 11: 145–148 - PubMed
-
- Afzal M., 1988. Consequences of consanguinity on cognitive behavior. Behav. Genet. 18: 583–594 - PubMed
-
- Bittles A. H., Black M. L., 2010. b The impact of consanguinity on neonatal and infant health. Early Hum. Dev. 86: 1779–1786 - PubMed
-
- Brown D. E., 1991. Human Universals. McGraw-Hill, New York
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
