

Beyond faithful conduction: short-term dynamics, neuromodulation, and long-term regulation of spike propagation in the axon
- PMID: 21708220
- PMCID: PMC3156869
- DOI: 10.1016/j.pneurobio.2011.06.001
Beyond faithful conduction: short-term dynamics, neuromodulation, and long-term regulation of spike propagation in the axon
Abstract
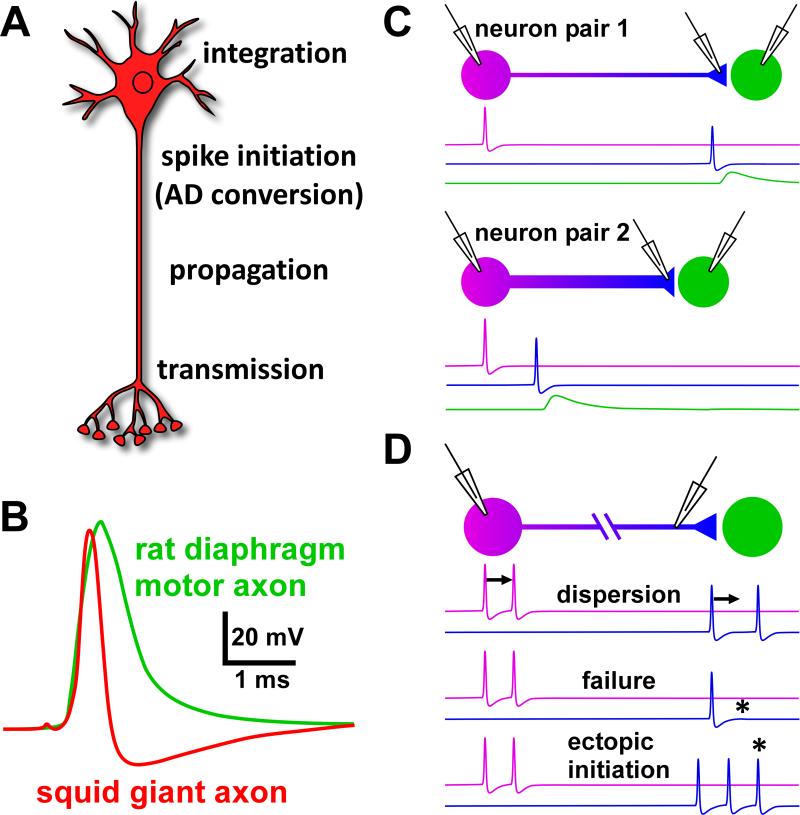
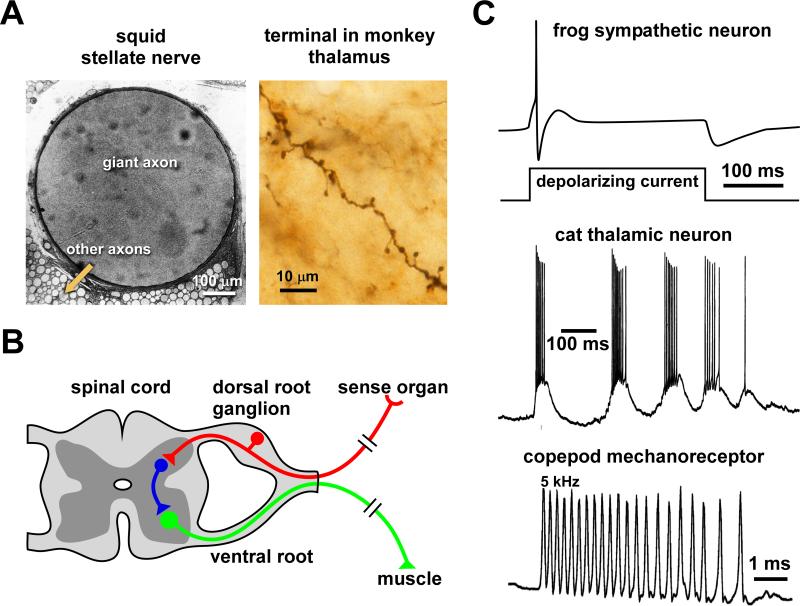
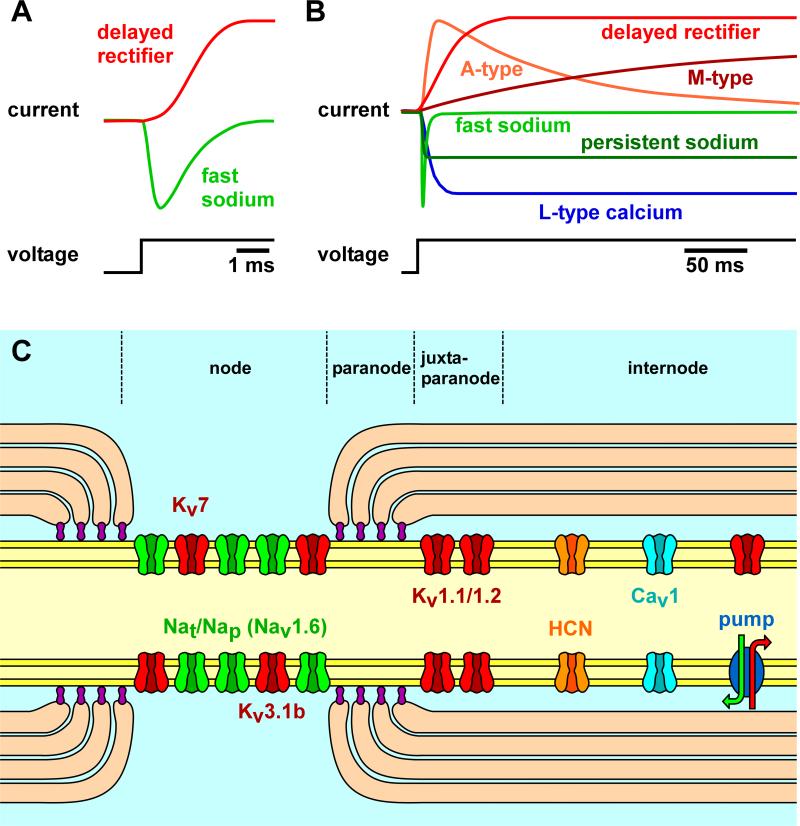
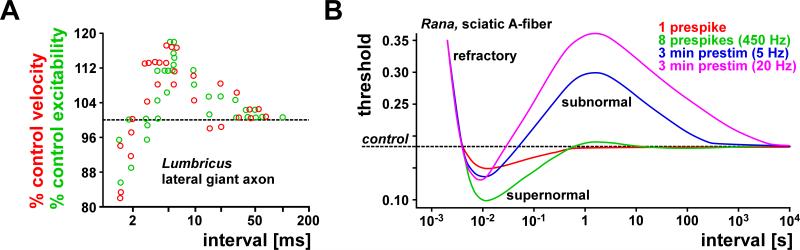
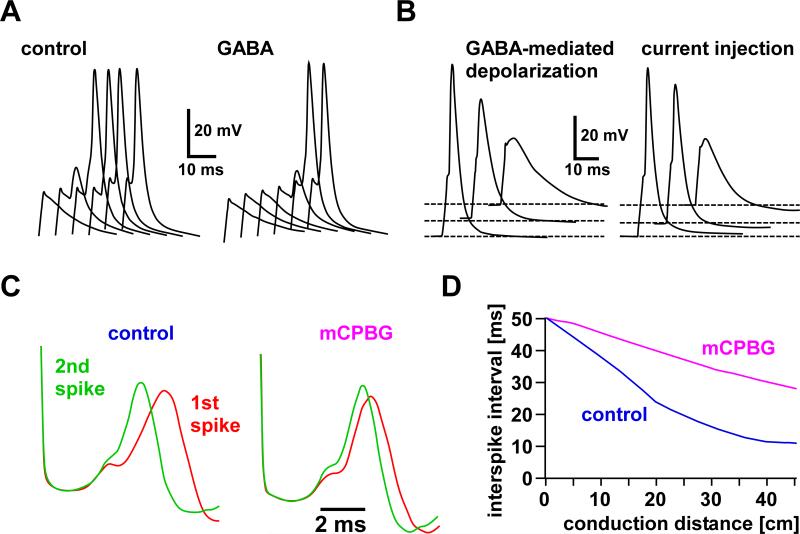
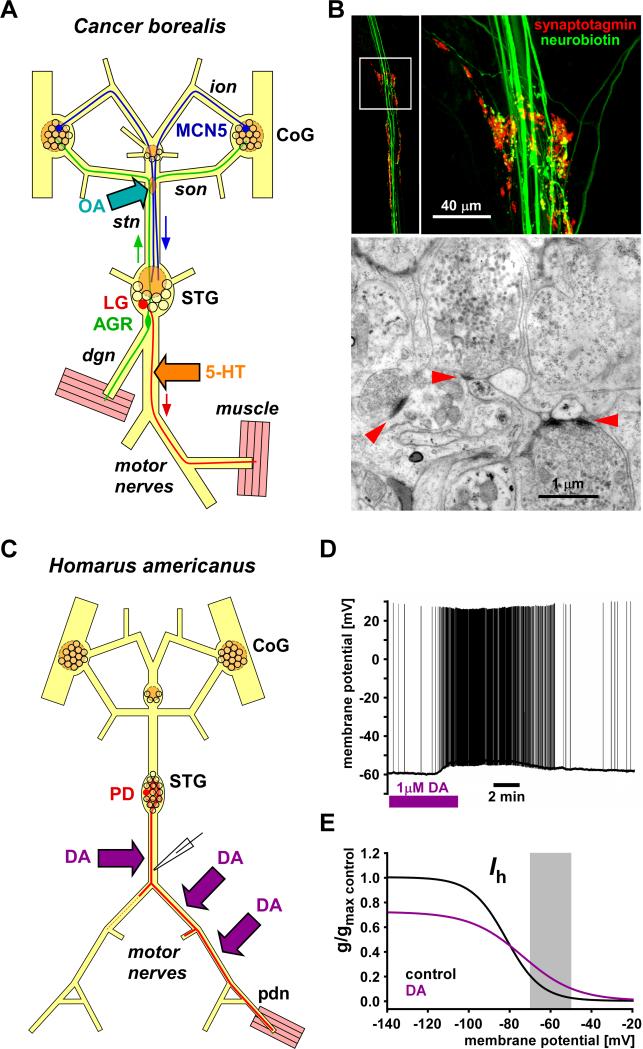
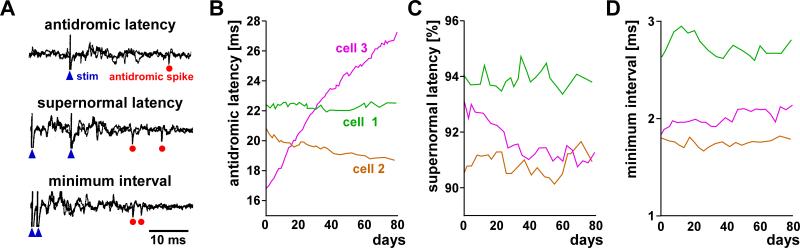
Most spiking neurons are divided into functional compartments: a dendritic input region, a soma, a site of action potential initiation, an axon trunk and its collaterals for propagation of action potentials, and distal arborizations and terminals carrying the output synapses. The axon trunk and lower order branches are probably the most neglected and are often assumed to do nothing more than faithfully conducting action potentials. Nevertheless, there are numerous reports of complex membrane properties in non-synaptic axonal regions, owing to the presence of a multitude of different ion channels. Many different types of sodium and potassium channels have been described in axons, as well as calcium transients and hyperpolarization-activated inward currents. The complex time- and voltage-dependence resulting from the properties of ion channels can lead to activity-dependent changes in spike shape and resting potential, affecting the temporal fidelity of spike conduction. Neural coding can be altered by activity-dependent changes in conduction velocity, spike failures, and ectopic spike initiation. This is true under normal physiological conditions, and relevant for a number of neuropathies that lead to abnormal excitability. In addition, a growing number of studies show that the axon trunk can express receptors to glutamate, GABA, acetylcholine or biogenic amines, changing the relative contribution of some channels to axonal excitability and therefore rendering the contribution of this compartment to neural coding conditional on the presence of neuromodulators. Long-term regulatory processes, both during development and in the context of activity-dependent plasticity may also affect axonal properties to an underappreciated extent.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
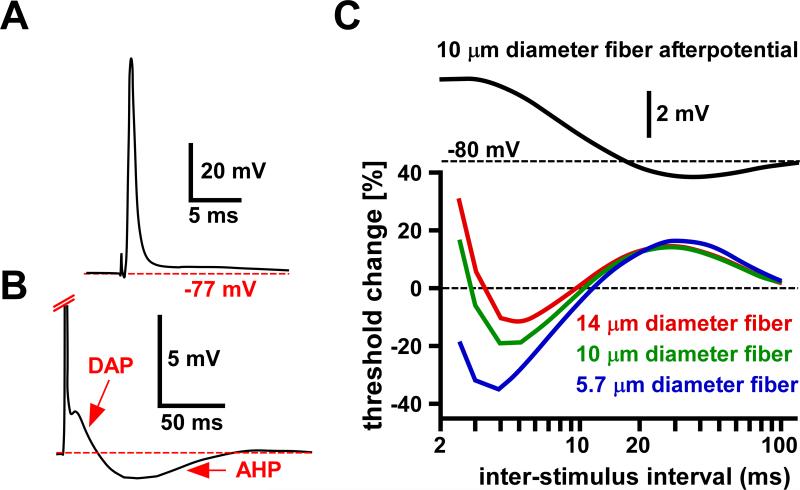
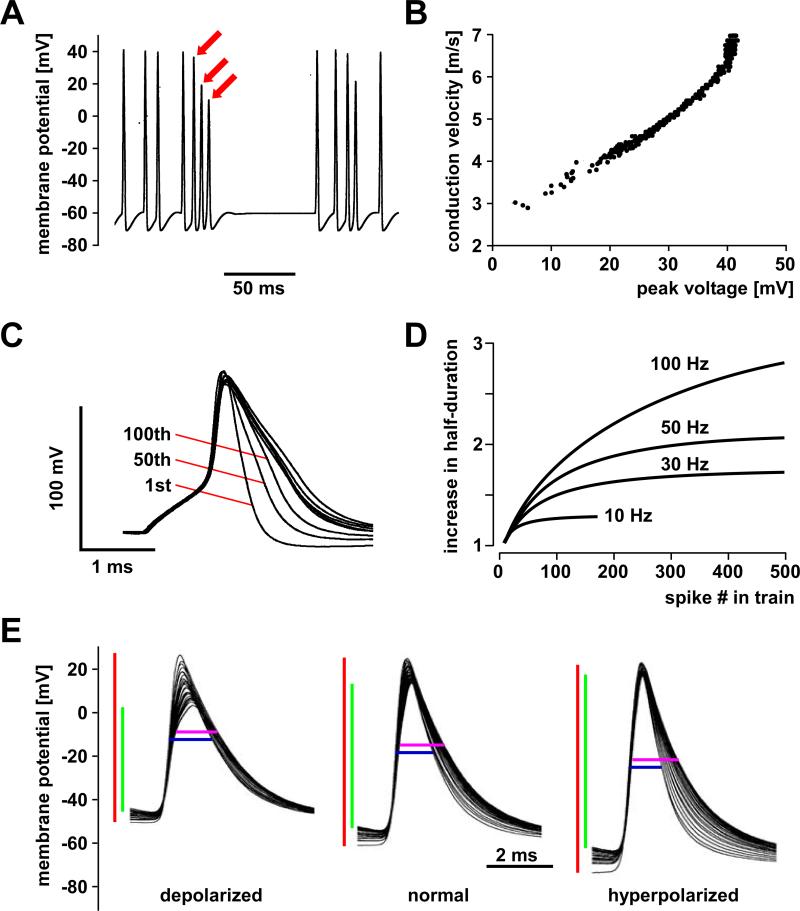
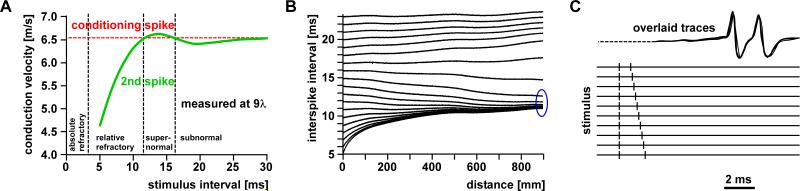
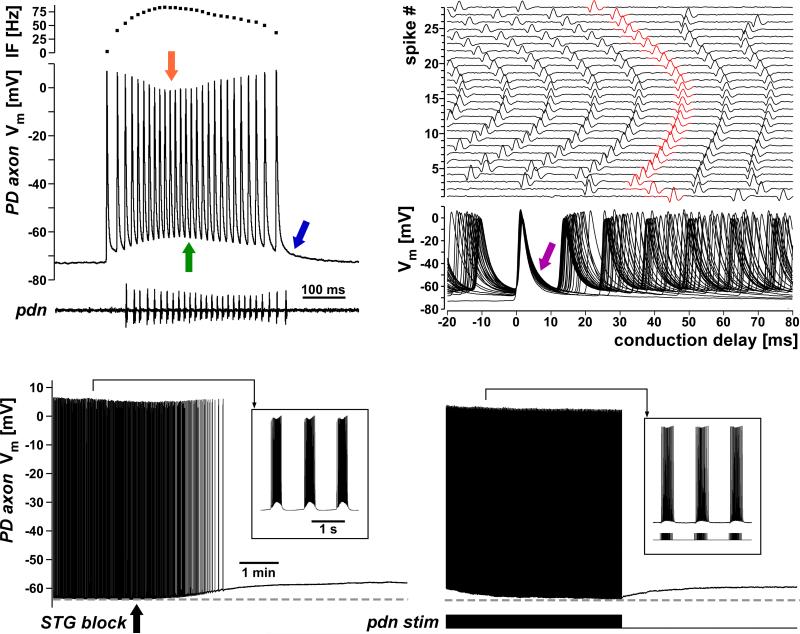
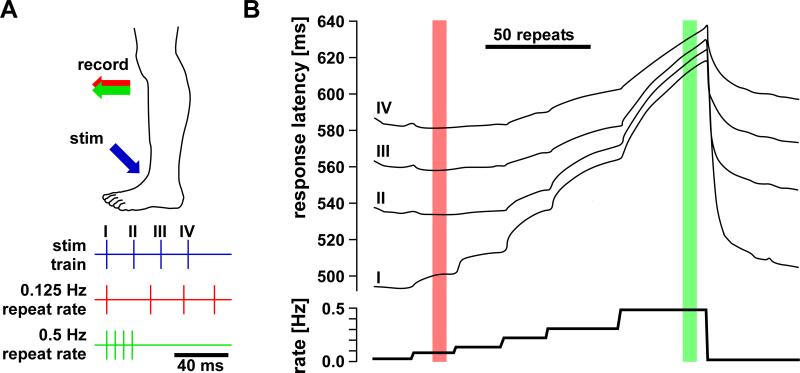
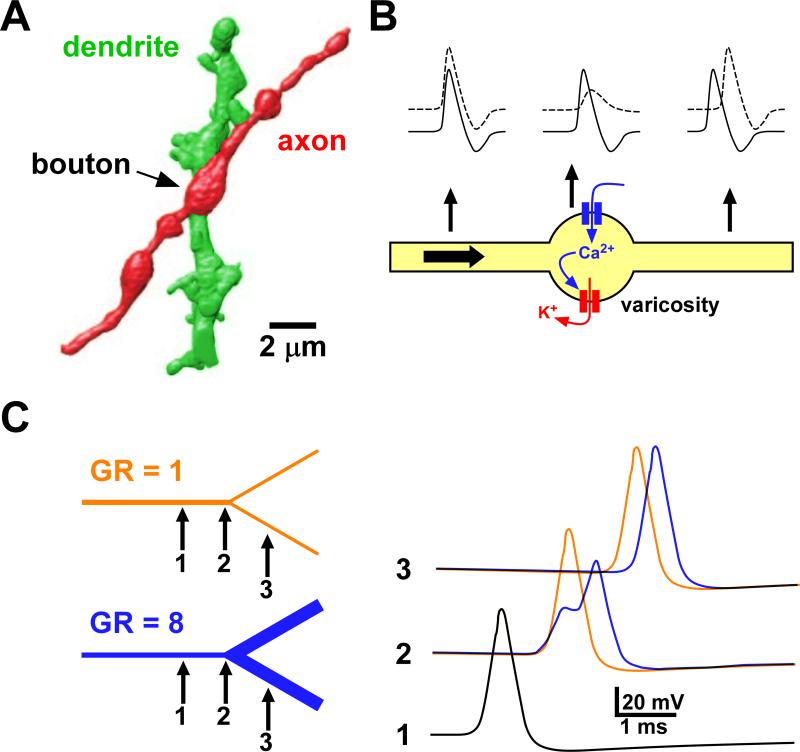
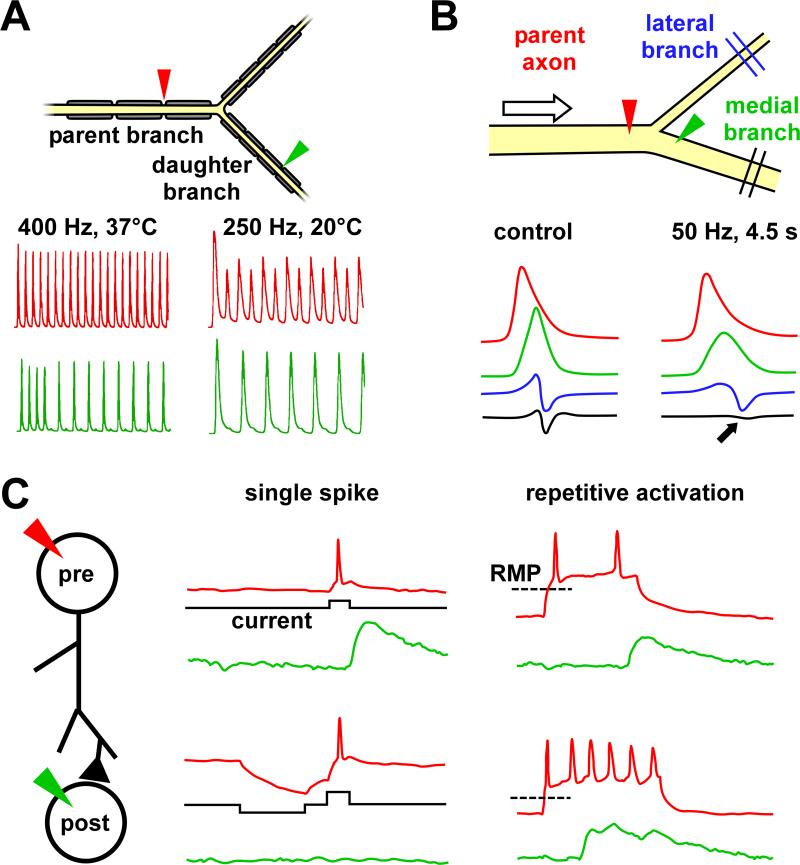
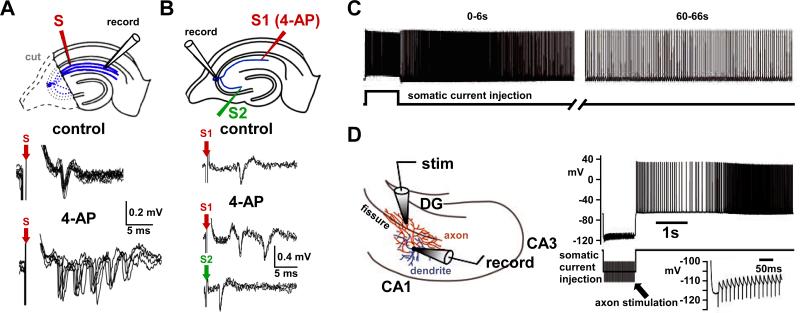
References
-
- Alle H, Geiger JR. Combined analog and action potential coding in hippocampal mossy fibers. Science. 2006;311:1290–1293. - PubMed
-
- Alle H, Geiger JR. Analog signalling in mammalian cortical axons. Curr Opin Neurobiol. 2008;18:314–320. - PubMed
-
- Alle H, Roth A, Geiger JR. Energy-efficient action potentials in hippocampal mossy fibers. Science. 2009;325:1405–1408. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
