

Review
doi: 10.1242/dmm.006791.
Zebrafish as a model for normal and malignant hematopoiesis
Affiliations
- PMID: 21708900
- PMCID: PMC3124047
- DOI: 10.1242/dmm.006791
Item in Clipboard
Review
Zebrafish as a model for normal and malignant hematopoiesis
Dis Model Mech.
2011 Jul.
Abstract
Zebrafish studies in the past two decades have made major contributions to our understanding of hematopoiesis and its associated disorders. The zebrafish has proven to be a powerful organism for studies in this area owing to its amenability to large-scale genetic and chemical screening. In addition, the externally fertilized and transparent embryos allow convenient genetic manipulation and in vivo imaging of normal and aberrant hematopoiesis. This review discusses available methods for studying hematopoiesis in zebrafish, summarizes key recent advances in this area, and highlights the current and potential contributions of zebrafish to the discovery and development of drugs to treat human blood disorders.
Figures
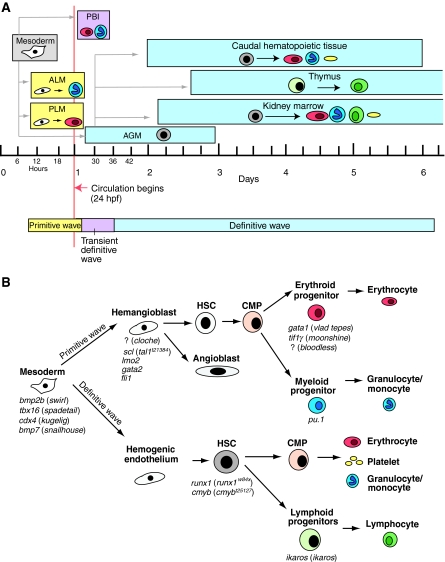
The ontogeny of hematopoiesis in zebrafish. (A) Hematopoiesis in zebrafish occurs in consecutive waves. Embryonic primitive hematopoiesis (yellow) starts at around 11 hours post fertilization (hpf) when hemangioblasts (which have the potential to become either endothelial vascular cells or hematopoietic cells, shown in white) appear in the anterior lateral mesoderm (ALM) and posterior lateral mesoderm (PLM), which collectively are analogous to the blood islands in the mammalian yolk sac. Hemangioblasts in the PLM later converge medially to form the intermediate cell mass (ICM; not shown), where primitive erythrocytes (red) predominantly arise. The ALM, which later becomes the rostral blood island (RBI; not shown), is the major site for primitive myeloid cells (blue). At around 24 hpf, embryonic erythrocytes enter circulation. As circulation begins, hematopoiesis within the ICM gradually diminishes. A transient definitive wave (pink) initiates shortly after multi-lineage erythromyeloid progenitors appear in the posterior blood island (PBI). Starting from 26 hpf, definitive HSCs (gray) emerge from hemogenic endothelial cells of the dorsal aorta in the aorta-gonad-mesonephros (AGM) region. Shortly thereafter, HSCs migrate to and seed the caudal hematopoietic tissue at 48 hpf, which is an expansion of the PBI and acts as a transient hematopoietic site that gives rise to erythroid, myeloid and thromboid (yellow) cells. The caudal hematopoietic tissue is equivalent to mouse fetal liver or placenta. HSCs from the AGM region colonize kidney around 48 hpf. Kidney marrow, which is functionally similar to mammalian bone marrow, gives rise to all blood lineages, including erythroid, myeloid, thromboid and lymphoid (green) cells for the larval and adult zebrafish. At around 54 hpf, common lymphoid progenitor (CLP) cells from the AGM region seed the thymus, which is the site for maturation of lymphoid T cells. (B) Crucial genes during different stages of hematopoietic development in zebrafish. The names of corresponding available zebrafish mutants are shown in parenthesis; the mutated genes are unknown for the mutants preceded by ‘?’. Adapted from Chen and Zon (Chen and Zon, 2009) and Paik and Zon (Paik and Zon, 2010), with permission.
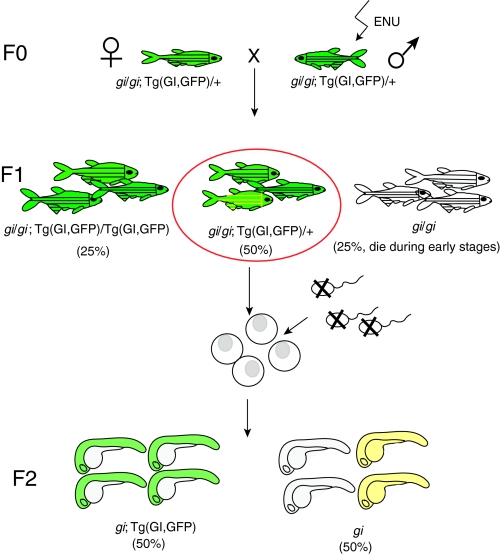
Scheme of a genetic suppressor screen. Lethal mutations in a gene of interest (gi) can be rescued by co-expressing a transgene in which a wild-type copy of the gene (GI) marked with GFP is driven by an appropriate promoter [note that a bacterial artificial chromosome (BAC) transgene that contains the entire locus of the GI gene could be used in the absence of suitable promoter, as in the case for moonshine (Bai et al., 2010)], enabling survival to adulthood (F0). Then, male F0 fish are mutagenized with the chemical ENU and are crossed to untreated female fish to generate F1 progeny. The F1 female fish that carry one copy of the transgene are used for subsequent in vitro fertilization. Eggs are squeezed out from the mother and fertilized with UV-treated sperm (which still activates the eggs but does not contribute chromosomes). All of the eggs develop into haploid embryos (F2), which stay alive until 5 days post-fertilization. 50% of the F2 embryos are GFP-positive (i.e. they express the rescuing transgene) and develop normally. The other 50% of haploid embryos are GFP-negative (i.e. they do not express the rescuing transgene) and develop recessive mutant phenotypes. However, if the mother carries a suppressor mutation in its genome (contributed by the ENU-mutagenized F0 male), 50% of the GFP-negative haploid embryos will be rescued (yellow).
References
-
- Bertrand J. Y., Traver D. (2009). Hematopoietic cell development in the zebrafish embryo. Curr. Opin. Hematol. 16, 243–248 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
