

Review
doi: 10.1083/jcb.201008173.
Evolution: structural and functional diversity of cadherin at the adherens junction
Affiliations
- PMID: 21708975
- PMCID: PMC3216324
- DOI: 10.1083/jcb.201008173
Item in Clipboard
Review
Evolution: structural and functional diversity of cadherin at the adherens junction
J Cell Biol.
.
Abstract
Adhesion between cells is essential to the evolution of multicellularity. Indeed, morphogenesis in animals requires firm but flexible intercellular adhesions that are mediated by subcellular structures like the adherens junction (AJ). A key component of AJs is classical cadherins, a group of transmembrane proteins that maintain dynamic cell-cell associations in many animal species. An evolutionary reconstruction of cadherin structure and function provides a comprehensive framework with which to appreciate the diversity of morphogenetic mechanisms in animals.
Figures
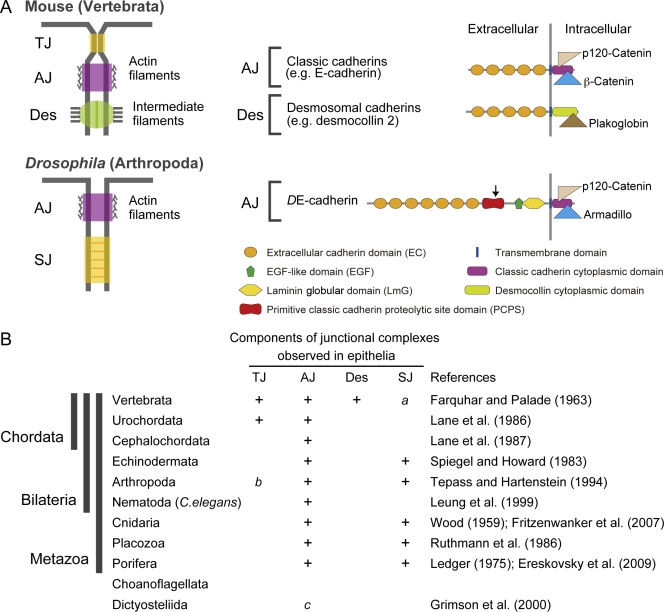
Diversity of intercellular junctions in the Metazoa. (A) Components of the vertebrate and Drosophila junctional complexes found in typical mature epithelia. TJ, tight junction; AJ, adherens junction; Des, desmosome; SJ, septate junction. Cadherins and associated proteins that constitute the AJ and the Des are shown. The arrow at the PCPS of DE-cadherin indicates the proteolytic cleavage site. (B) Phylogenetic distributions of the junction types observed in epithelia. “a” denotes that SJ-like junctions are present in the nervous system of vertebrates (Bellen et al., 1998); “b” denotes that TJ-like junctions are present in the nervous system of chelicerate arthropods (Lane, 2001); and “c” denotes that AJ-like junctions are present in the slime mold Dictyostelium References: Wood, 1959; Farquhar and Palade, 1963; Ledger, 1975; Spiegel and Howard, 1983; Lane et al., 1986, ; Ruthmann et al., 1986; Tepass and Hartenstein, 1994; Leung et al., 1999; Grimson et al., 2000; Fritzenwanker et al., 2007; Ereskovsky et al., 2009.
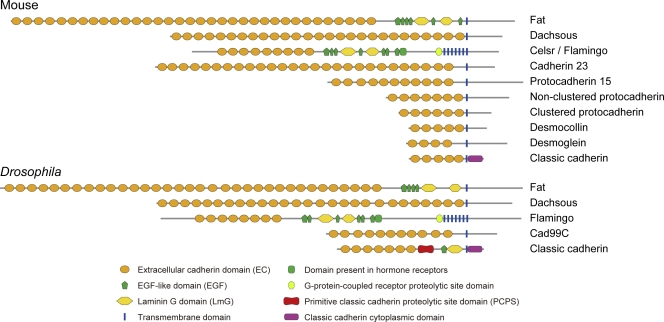
The domain structures of representative members of the major cadherin subfamilies in the mouse and Drosophila. Shown is a comparison of the domain structures based on predictions of domains by SMART/Pfam analysis, except for the PCPS in DE-cadherin. Some cadherin subfamilies are missing in Drosophila.
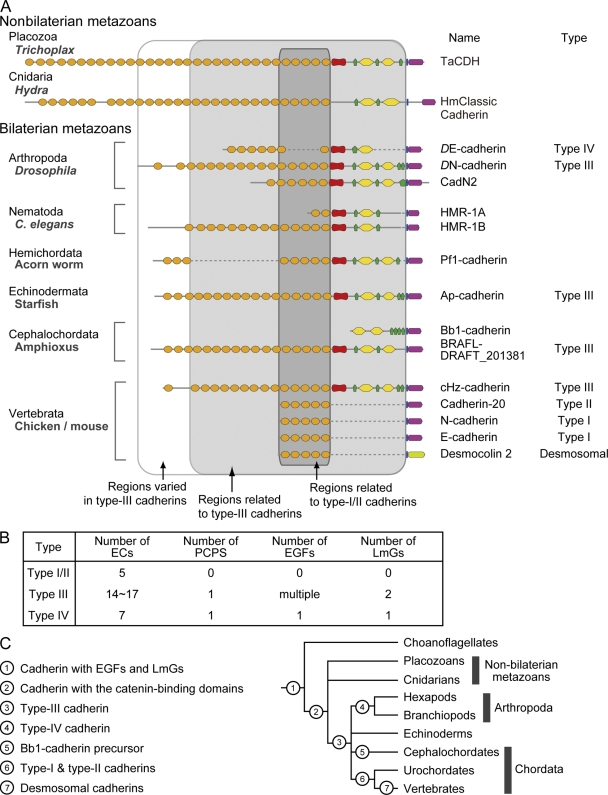
Diversity and evolution of the extracellular domain structures of classical cadherins in the Metazoa. (A) Comparison of the domain structures of selected classical cadherins and a desmosomal cadherin in bilaterian and nonbilaterian metazoan phyla. Gaps (dotted lines) are introduced to highlight homologous extracellular regions between distinct cadherins. (B) Comparison of the domain compositions among the classical cadherin types. (C) A possible phylogenetic diagram showing key genomic changes that contributed to the structural diversification of cadherins at the AJ and the desmosome.
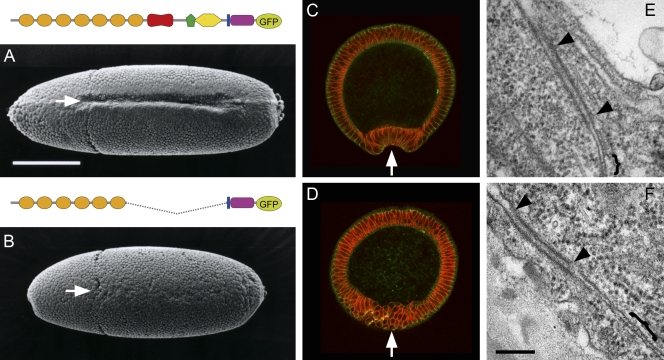
Replacement of DE-cadherin with a shortened DE-cadherin, DEΔP. Shown are Drosophila embryos in which DE-cadherin was replaced with DE-cadherin-GFP (A, C, and E) or DEΔP (B, D, and F), which was also tagged with GFP. (A and B) Scanning electron microscopy images showing the ventral view of early gastrulation–stage embryos. (C and D) Cross sections of early gastrulation–stage embryos. Green, GFP fluorescence; red, neurotactin staining. The ventral furrow is seen in A and C but not in B and D (arrows). The ectodermal epithelia developed normally in both embryo types. (E and F) Transmission electron microscopy images showing an apical region of cell contact in the lateral epidermis of late-stage embryos. AJ (arrowheads) and SJ (brackets) are seen in both E and F, where there are no recognizable distinctions. Bars: (A) 200 µm; (F) 200 nm. Adapted from Haruta et al., 2010 with permission from John Wiley and Sons.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
