

The p25 subunit of the dynactin complex is required for dynein-early endosome interaction
- PMID: 21708978
- PMCID: PMC3216330
- DOI: 10.1083/jcb.201011022
The p25 subunit of the dynactin complex is required for dynein-early endosome interaction
Abstract
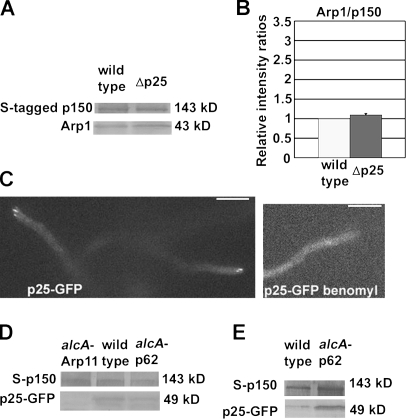
Cytoplasmic dynein transports various cellular cargoes including early endosomes, but how dynein is linked to early endosomes is unclear. We find that the Aspergillus nidulans orthologue of the p25 subunit of dynactin is critical for dynein-mediated early endosome movement but not for dynein-mediated nuclear distribution. In the absence of NUDF/LIS1, p25 deletion abolished the localization of dynein-dynactin to the hyphal tip where early endosomes abnormally accumulate but did not prevent dynein-dynactin localization to microtubule plus ends. Within the dynactin complex, p25 locates at the pointed end of the Arp1 filament with Arp11 and p62, and our data suggest that Arp11 but not p62 is important for p25-dynactin association. Loss of either Arp1 or p25 significantly weakened the physical interaction between dynein and early endosomes, although loss of p25 did not apparently affect the integrity of the Arp1 filament. These results indicate that p25, in conjunction with the rest of the dynactin complex, is important for dynein-early endosome interaction.
Figures
Similar articles
-
Cargo-Mediated Activation of Cytoplasmic Dynein in vivo.Front Cell Dev Biol. 2020 Oct 23;8:598952. doi: 10.3389/fcell.2020.598952. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33195284 Free PMC article. Review.
-
p25 of the dynactin complex plays a dual role in cargo binding and dynactin regulation.J Biol Chem. 2018 Oct 5;293(40):15606-15619. doi: 10.1074/jbc.RA118.004000. Epub 2018 Aug 24. J Biol Chem. 2018. PMID: 30143531 Free PMC article.
-
HookA is a novel dynein-early endosome linker critical for cargo movement in vivo.J Cell Biol. 2014 Mar 17;204(6):1009-26. doi: 10.1083/jcb.201308009. J Cell Biol. 2014. PMID: 24637327 Free PMC article.
-
The actin capping protein in Aspergillus nidulans enhances dynein function without significantly affecting Arp1 filament assembly.Sci Rep. 2018 Jul 30;8(1):11419. doi: 10.1038/s41598-018-29818-4. Sci Rep. 2018. PMID: 30061726 Free PMC article.
-
Cytoplasmic dynein and early endosome transport.Cell Mol Life Sci. 2015 Sep;72(17):3267-80. doi: 10.1007/s00018-015-1926-y. Epub 2015 May 23. Cell Mol Life Sci. 2015. PMID: 26001903 Free PMC article. Review.
Cited by
-
Cargo-Mediated Activation of Cytoplasmic Dynein in vivo.Front Cell Dev Biol. 2020 Oct 23;8:598952. doi: 10.3389/fcell.2020.598952. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33195284 Free PMC article. Review.
-
VezA/vezatin facilitates proper assembly of the dynactin complex in vivo.Cell Rep. 2024 Nov 26;43(11):114943. doi: 10.1016/j.celrep.2024.114943. Epub 2024 Nov 1. Cell Rep. 2024. PMID: 39487986 Free PMC article.
-
Secretory Vesicle Polar Sorting, Endosome Recycling and Cytoskeleton Organization Require the AP-1 Complex in Aspergillus nidulans.Genetics. 2018 Aug;209(4):1121-1138. doi: 10.1534/genetics.118.301240. Epub 2018 Jun 20. Genetics. 2018. PMID: 29925567 Free PMC article.
-
VezA/vezatin facilitates proper assembly of the dynactin complex in vivo.bioRxiv [Preprint]. 2024 Apr 20:2024.04.19.590248. doi: 10.1101/2024.04.19.590248. bioRxiv. 2024. Update in: Cell Rep. 2024 Nov 26;43(11):114943. doi: 10.1016/j.celrep.2024.114943. PMID: 38659795 Free PMC article. Updated. Preprint.
-
Cryo-EM reveals the complex architecture of dynactin's shoulder region and pointed end.EMBO J. 2021 Apr 15;40(8):e106164. doi: 10.15252/embj.2020106164. Epub 2021 Mar 18. EMBO J. 2021. PMID: 33734450 Free PMC article.
References
-
- Borkovich K.A., Alex L.A., Yarden O., Freitag M., Turner G.E., Read N.D., Seiler S., Bell-Pedersen D., Paietta J., Plesofsky N., et al. 2004. Lessons from the genome sequence of Neurospora crassa: tracing the path from genomic blueprint to multicellular organism. Microbiol. Mol. Biol. Rev. 68:1–108 10.1128/MMBR.68.1.1-108.2004 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
