

The cytokinin receptors of Arabidopsis are located mainly to the endoplasmic reticulum
- PMID: 21709172
- PMCID: PMC3149959
- DOI: 10.1104/pp.111.180539
The cytokinin receptors of Arabidopsis are located mainly to the endoplasmic reticulum
Abstract
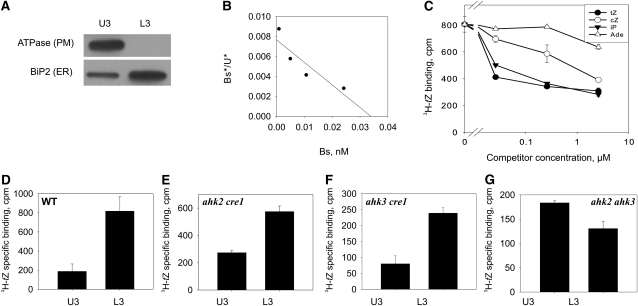
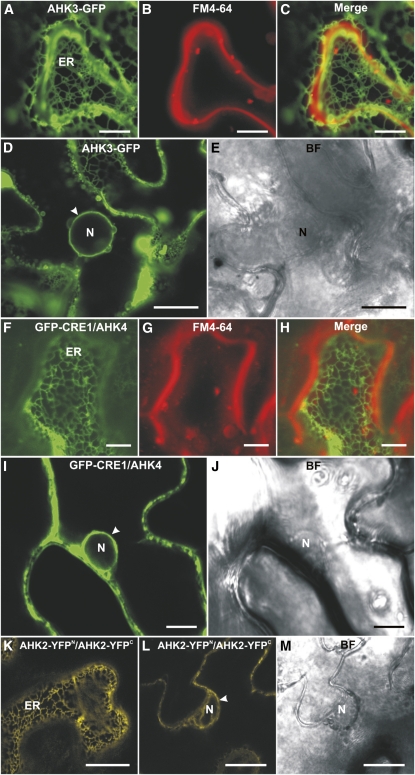
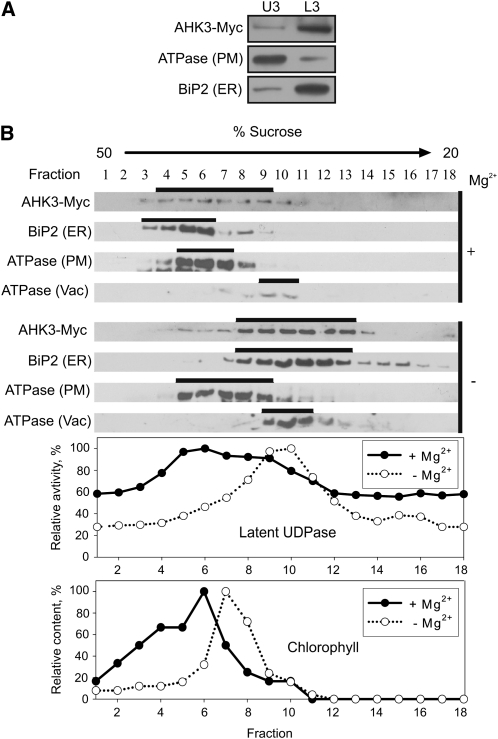
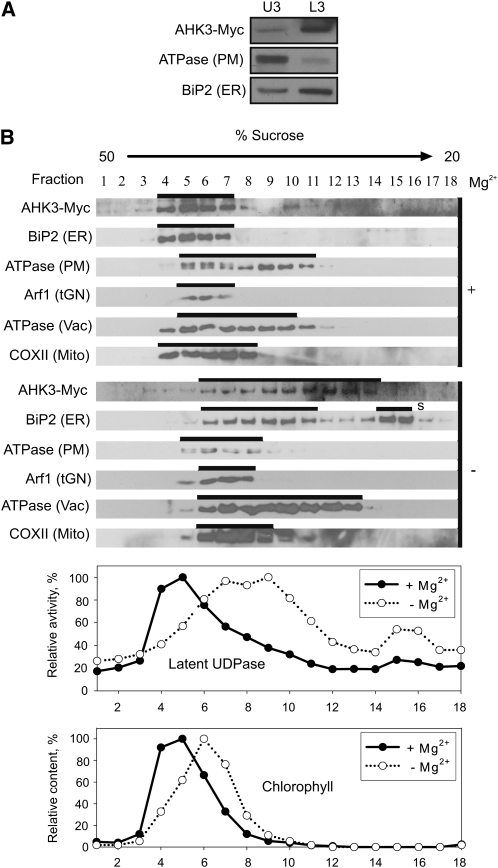
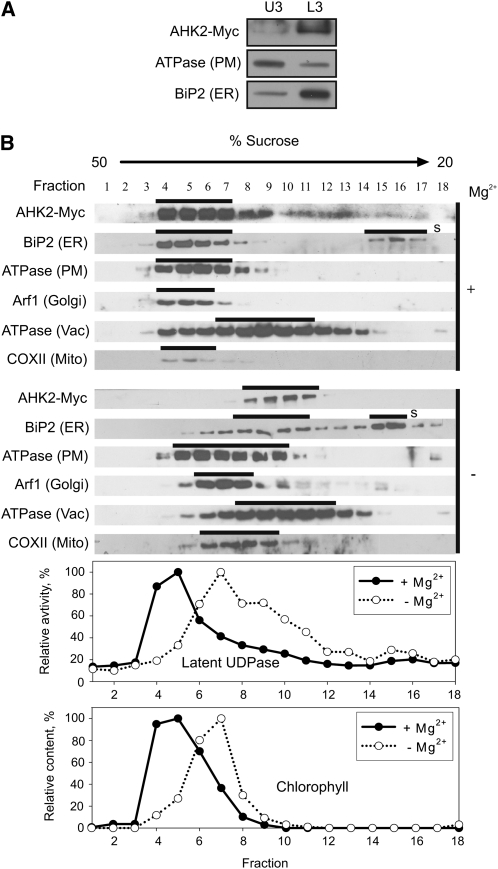
The plant hormone cytokinin is perceived by membrane-located sensor histidine kinases. Arabidopsis (Arabidopsis thaliana) possesses three cytokinin receptors: ARABIDOPSIS HISTIDINE KINASE2 (AHK2), AHK3, and CYTOKININ RESPONSE1/AHK4. The current model predicts perception of the cytokinin signal at the plasma membrane. However, cytokinin-binding studies with membrane fractions separated by two-phase partitioning showed that in the wild type, as well as in mutants retaining only single cytokinin receptors, the major part of specific cytokinin binding was associated with endomembranes. Leaf epidermal cells of tobacco (Nicotiana benthamiana) expressing receptor-green fluorescent protein fusion proteins and bimolecular fluorescence complementation analysis showed strong fluorescence of the endoplasmic reticulum (ER) network for all three receptors. Furthermore, separation of the microsomal fraction of Arabidopsis plants expressing Myc-tagged AHK2 and AHK3 receptors by sucrose gradient centrifugation followed by immunoblotting displayed the Mg²⁺-dependent density shift typical of ER membrane proteins. Cytokinin-binding assays, fluorescent fusion proteins, and biochemical fractionation all showed that the large majority of cytokinin receptors are localized to the ER, suggesting a central role of this compartment in cytokinin signaling. A modified model for cytokinin signaling is proposed.
Figures
References
-
- Argueso CT, Ferreira FJ, Kieber JJ. (2009) Environmental perception avenues: the interaction of cytokinin and environmental response pathways. Plant Cell Environ 32: 1147–1160 - PubMed
-
- Bishopp A, Mähönen AP, Helariutta Y. (2006) Signs of change: hormone receptors that regulate plant development. Development 133: 1857–1869 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
