

Taste-independent detection of the caloric content of sugar in Drosophila
- PMID: 21709242
- PMCID: PMC3136275
- DOI: 10.1073/pnas.1017096108
Taste-independent detection of the caloric content of sugar in Drosophila
Abstract
Feeding behavior is influenced primarily by two factors: nutritional needs and food palatability. However, the role of food deprivation and metabolic needs in the selection of appropriate food is poorly understood. Here, we show that the fruit fly, Drosophila melanogaster, selects calorie-rich foods following prolonged food deprivation in the absence of taste-receptor signaling. Flies mutant for the sugar receptors Gr5a and Gr64a cannot detect the taste of sugar, but still consumed sugar over plain agar after 15 h of starvation. Similarly, pox-neuro mutants that are insensitive to the taste of sugar preferentially consumed sugar over plain agar upon starvation. Moreover, when given a choice between metabolizable sugar (sucrose or D-glucose) and nonmetabolizable (zero-calorie) sugar (sucralose or L-glucose), starved Gr5a; Gr64a double mutants preferred metabolizable sugars. These findings suggest the existence of a taste-independent metabolic sensor that functions in food selection. The preference for calorie-rich food correlates with a decrease in the two main hemolymph sugars, trehalose and glucose, and in glycogen stores, indicating that this sensor is triggered when the internal energy sources are depleted. Thus, the need to replenish depleted energy stores during periods of starvation may be met through the activity of a taste-independent metabolic sensing pathway.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
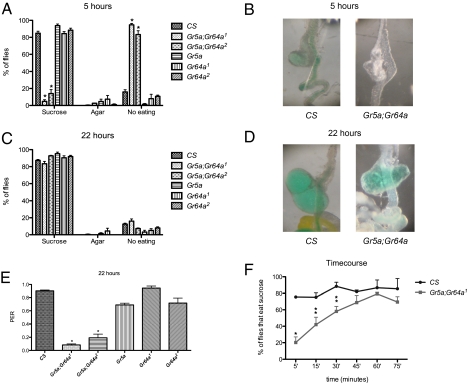

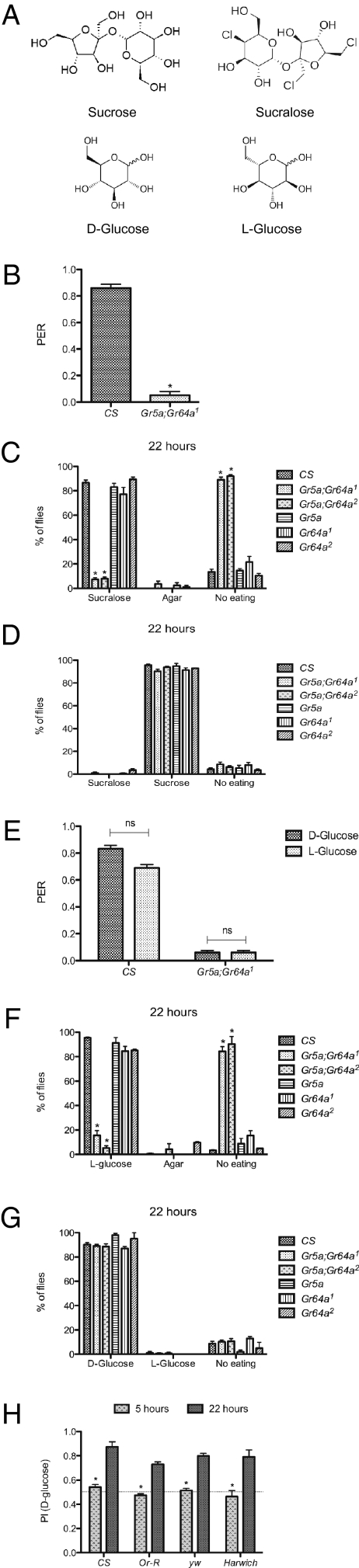

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
