

Pathway for lipid A biosynthesis in Arabidopsis thaliana resembling that of Escherichia coli
- PMID: 21709257
- PMCID: PMC3136250
- DOI: 10.1073/pnas.1108840108
Pathway for lipid A biosynthesis in Arabidopsis thaliana resembling that of Escherichia coli
Abstract
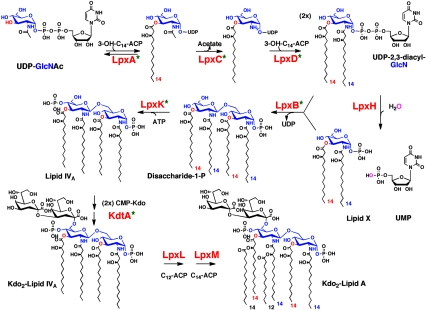
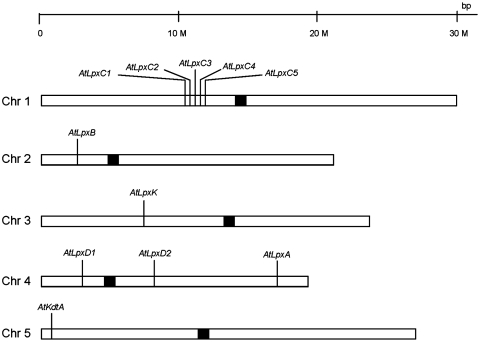
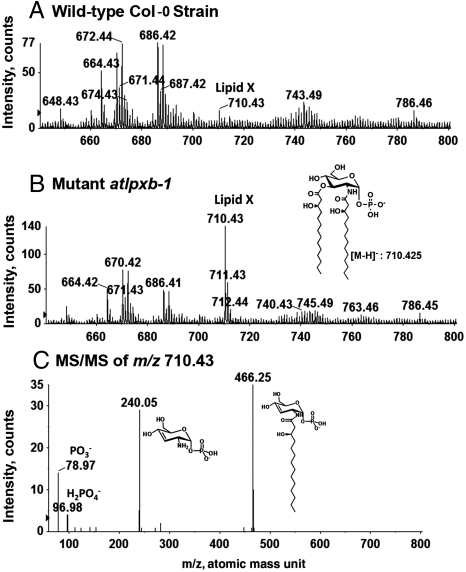
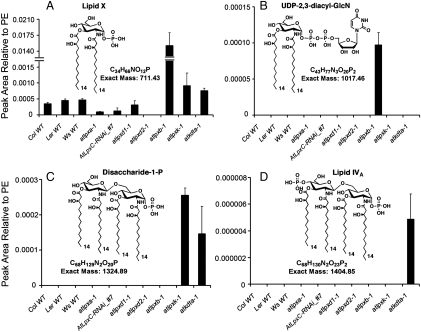
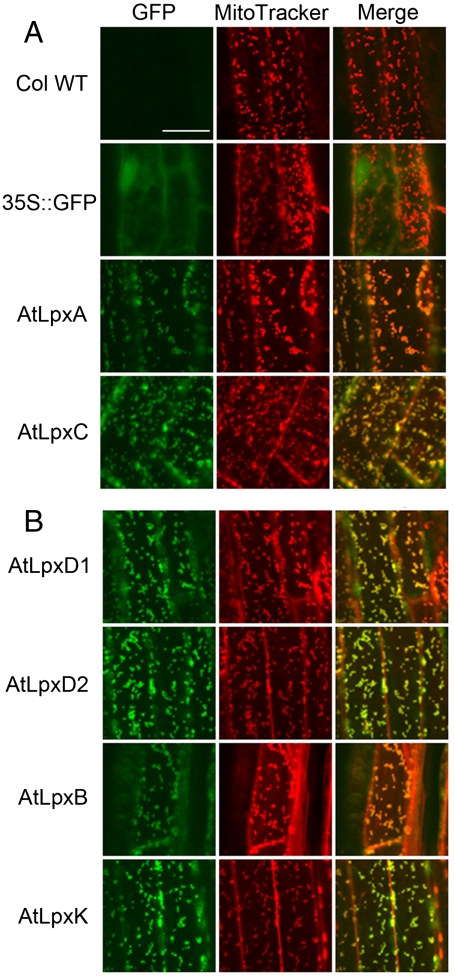
The lipid A moiety of Escherichia coli lipopolysaccharide is a hexa-acylated disaccharide of glucosamine that makes up the outer monolayer of the outer membrane. Arabidopsis thaliana contains nuclear genes encoding orthologs of key enzymes of bacterial lipid A biosynthesis, including LpxA, LpxC, LpxD, LpxB, LpxK and KdtA. Although structurally related lipid A molecules are found in most other gram-negative bacteria, lipid A and its precursors have not been directly detected in plants previously. However, homozygous insertional knockout mutations or RNAi knock-down constructs of Arabidopsis lpx and kdtA mutants revealed accumulation (or disappearance) of the expected monosaccharide or disaccharide lipid A precursors by mass spectrometry of total lipids extracted from 10-day old seedlings of these mutants. In addition, fluorescence microscopy of lpx-gfp fusions in transgenic Arabidopsis plants suggests that the Lpx and KdtA proteins are expressed and targeted to mitochondria. Although the structure of the lipid A end product generated by plants is still unknown, our work demonstrates that plants synthesize lipid A precursors using the same enzymatic pathway present in E. coli.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Activity and crystal structure of Arabidopsis thaliana UDP-N-acetylglucosamine acyltransferase.Biochemistry. 2012 May 29;51(21):4322-30. doi: 10.1021/bi3002242. Epub 2012 May 14. Biochemistry. 2012. PMID: 22545860 Free PMC article.
-
A Complete Pathway Model for Lipid A Biosynthesis in Escherichia coli.PLoS One. 2015 Apr 28;10(4):e0121216. doi: 10.1371/journal.pone.0121216. eCollection 2014. PLoS One. 2015. PMID: 25919634 Free PMC article.
-
Outer membrane permeability barrier in Escherichia coli mutants that are defective in the late acyltransferases of lipid A biosynthesis.Antimicrob Agents Chemother. 1999 Jun;43(6):1459-62. doi: 10.1128/AAC.43.6.1459. Antimicrob Agents Chemother. 1999. PMID: 10348770 Free PMC article.
-
The biosynthesis of gram-negative endotoxin. Formation of lipid A disaccharides from monosaccharide precursors in extracts of Escherichia coli.J Biol Chem. 1984 Apr 25;259(8):4852-9. J Biol Chem. 1984. PMID: 6370995
-
yciM is an essential gene required for regulation of lipopolysaccharide synthesis in Escherichia coli.Mol Microbiol. 2014 Jan;91(1):145-57. doi: 10.1111/mmi.12452. Epub 2013 Nov 24. Mol Microbiol. 2014. PMID: 24266962
Cited by
-
Poplar carbohydrate-active enzymes: whole-genome annotation and functional analyses based on RNA expression data.Plant J. 2019 Aug;99(4):589-609. doi: 10.1111/tpj.14417. Epub 2019 Jul 1. Plant J. 2019. PMID: 31111606 Free PMC article.
-
MAMP (microbe-associated molecular pattern) triggered immunity in plants.Front Plant Sci. 2013 May 16;4:139. doi: 10.3389/fpls.2013.00139. eCollection 2013. Front Plant Sci. 2013. PMID: 23720666 Free PMC article.
-
Activity and crystal structure of Arabidopsis thaliana UDP-N-acetylglucosamine acyltransferase.Biochemistry. 2012 May 29;51(21):4322-30. doi: 10.1021/bi3002242. Epub 2012 May 14. Biochemistry. 2012. PMID: 22545860 Free PMC article.
-
Acyl-lipid metabolism.Arabidopsis Book. 2013;11:e0161. doi: 10.1199/tab.0161. Epub 2013 Jan 29. Arabidopsis Book. 2013. PMID: 23505340 Free PMC article.
-
Immune Sensing of Lipopolysaccharide in Plants and Animals: Same but Different.PLoS Pathog. 2016 Jun 9;12(6):e1005596. doi: 10.1371/journal.ppat.1005596. eCollection 2016 Jun. PLoS Pathog. 2016. PMID: 27281177 Free PMC article. Review. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
