

Locostatin Disrupts Association of Raf Kinase Inhibitor Protein With Binding Proteins by Modifying a Conserved Histidine Residue in the Ligand-Binding Pocket
- PMID: 21709760
- PMCID: PMC3121171
- DOI: 10.1615/forumimmundisther.v2.i1.60
Locostatin Disrupts Association of Raf Kinase Inhibitor Protein With Binding Proteins by Modifying a Conserved Histidine Residue in the Ligand-Binding Pocket
Abstract
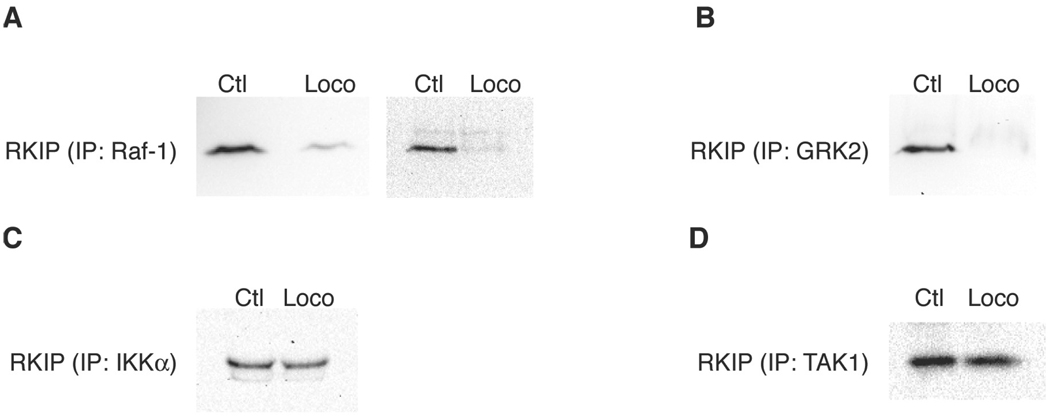
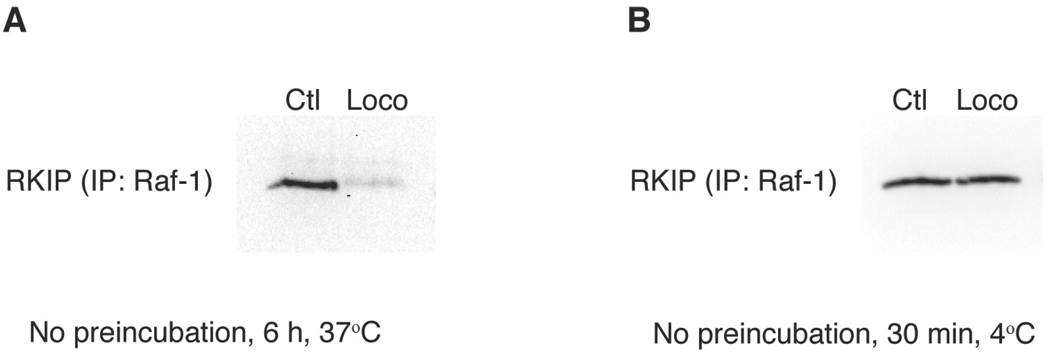
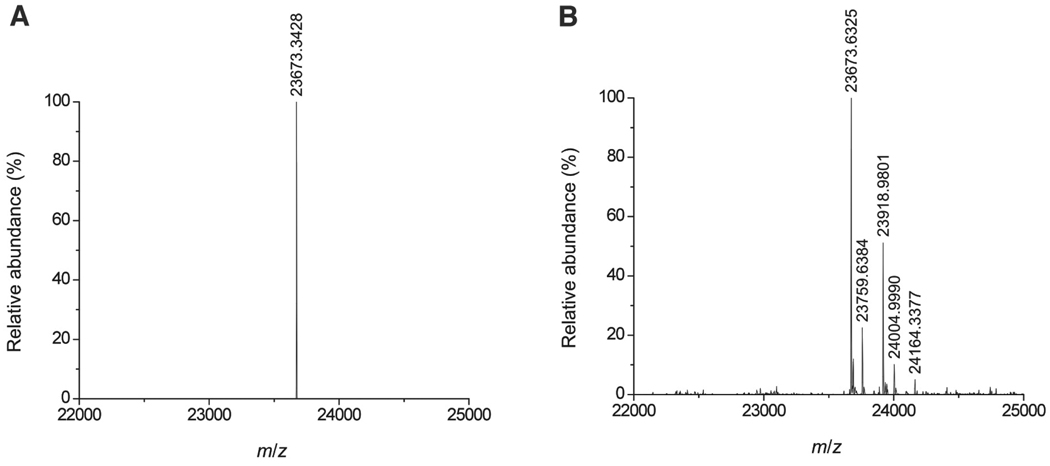
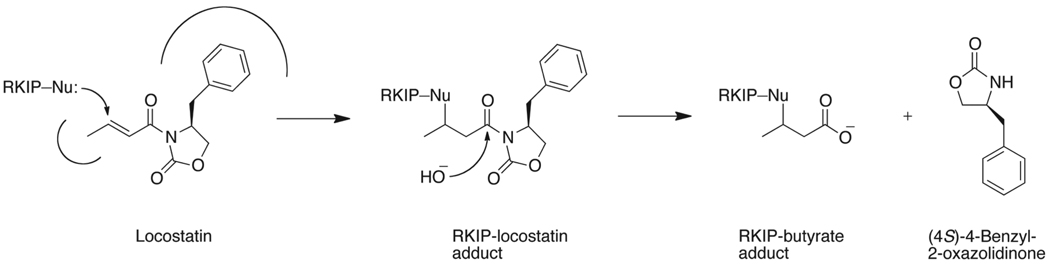
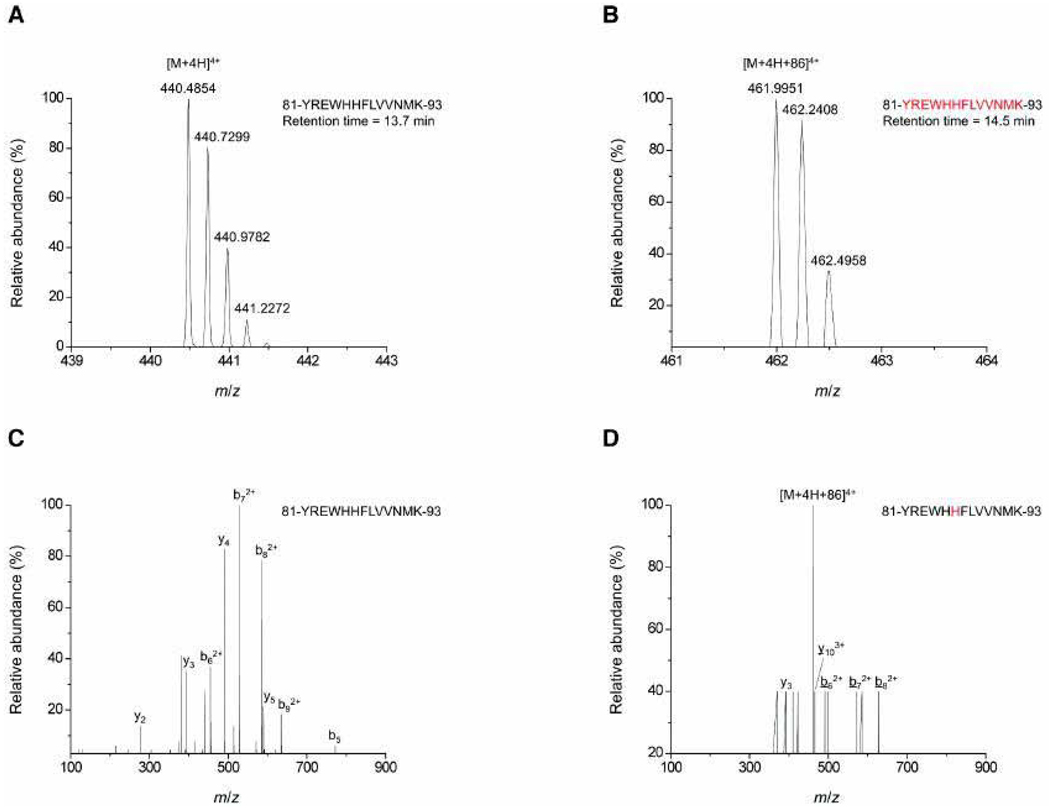
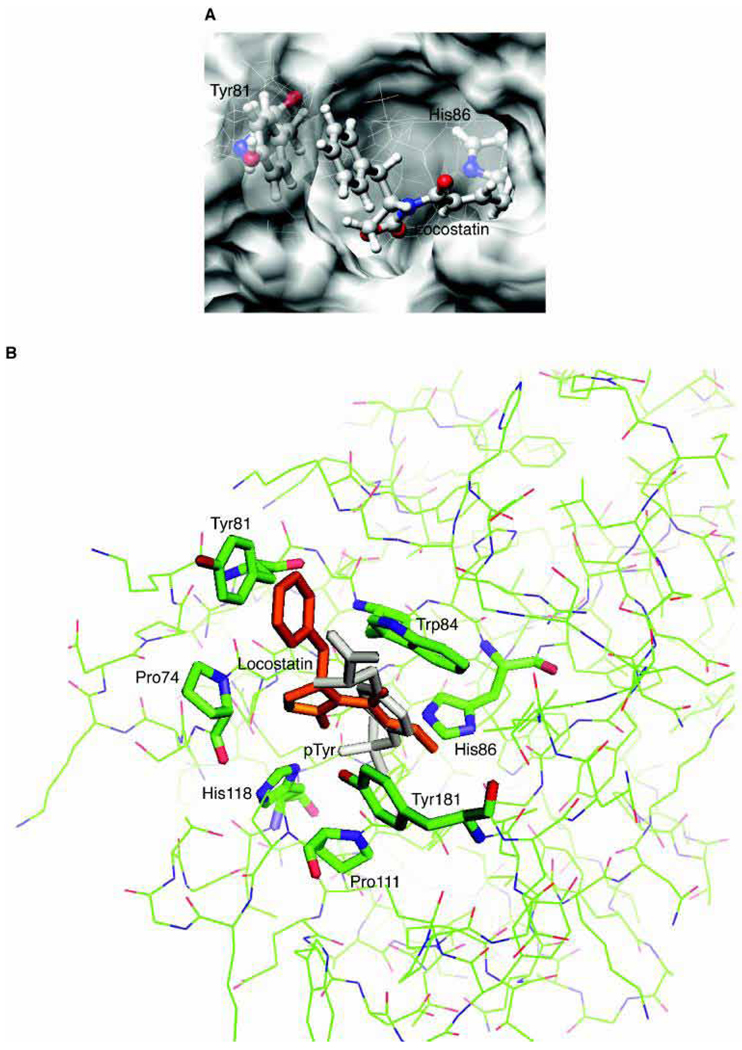
Raf kinase inhibitor protein (RKIP) interacts with a number of different proteins and regulates multiple signaling pathways. Here, we show that locostatin, a small molecule that covalently binds RKIP, not only disrupts interactions of RKIP with Raf-1 kinase, but also with G protein-coupled receptor kinase 2. In contrast, we found that locostatin does not disrupt binding of RKIP to two other proteins: inhibitor of κB kinase α and transforming growth factor β-activated kinase 1. These results thus imply that different proteins interact with different regions of RKIP. Locostatin's mechanism of action involves modification of a nucleophilic residue on RKIP. We observed that after binding RKIP, part of locostatin is slowly hydrolyzed, leaving a smaller RKIP-butyrate adduct. We identified the residue alkylated by locostatin as His86, a highly conserved residue in RKIP's ligand-binding pocket. Computational modeling of the binding of locostatin to RKIP suggested that the recognition interaction between small molecule and protein ensures that locostatin's electrophilic site is poised to react with His86. Furthermore, binding of locostatin would sterically hinder binding of other ligands in the pocket. These data provide a basis for understanding how locostatin disrupts particular interactions of RKIP with RKIP-binding proteins and demonstrate its utility as a probe of specific RKIP interactions and functions.
Figures
References
-
- Mc Henry KT, Ankala SV, Ghosh AK, Fenteany G. A non-antibacterial oxazolidinone derivative that inhibits epithelial cell sheet migration. ChemBioChem. 2002;3:1105–1111. - PubMed
-
- Mc Henry KT, Montesano R, Zhu S, Beshir AB, Tang HH, Yeung KC, Fenteany G. Raf kinase inhibitor protein positively regulates cell-substratum adhesion while negatively regulating cell-cell adhesion. J Cell Biochem. 2008;103:972–985. - PubMed
-
- Zhu S, Mc Henry KT, Lane WS, Fenteany G. A chemical inhibitor reveals the role of Raf kinase inhibitor protein in cell migration. Chem Biol. 2005;12:981–991. - PubMed
-
- Yeung K, Seitz T, Li S, Janosch P, McFerran B, Kaiser C, Fee F, Katsanakis KD, Rose DW, Mischak H, Sedivy JM, Kolch W. Suppression of Raf-1 kinase activity and MAP kinase signalling by RKIP. Nature. 1999;401:173–177. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous