

doi: 10.1167/iovs.10-7083.
Cell populations of the retina: the Proctor lecture
Affiliations
- PMID: 21712414
- PMCID: PMC3175966
- DOI: 10.1167/iovs.10-7083
Item in Clipboard
Cell populations of the retina: the Proctor lecture
Invest Ophthalmol Vis Sci.
.
No abstract available
Figures
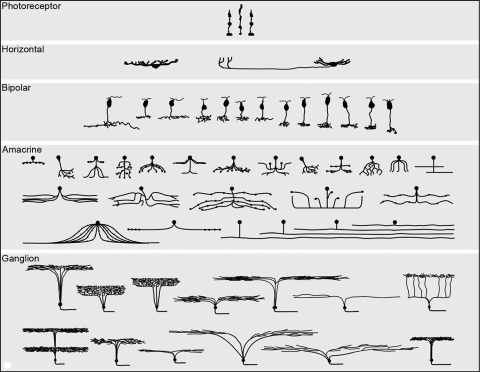
The major cell populations of a generic mammalian retina, shown semi-schematically in section view. Mammalian retinas contain in excess of 60 distinct cell types, each carrying out a distinct, dedicated function. The populations of photoreceptor, horizontal, and bipolar cells are represented quite completely. Amacrine cells are fairly complete, except that more types of wide-field amacrine cells exist than could be represented here. The ganglion cells shown are from a survey carried out in the retina of the rabbit. Ganglion cells vary more from species to species than other retinal cell types and there is reason to believe that, in most species, the number of ganglion cell types is at least 20. The illustration is reproduced with permission from Masland RH. The fundamental plan of the retina. Nature Neurosci. 2001;4:877–886, © Elsevier, where it is discussed in more detail.
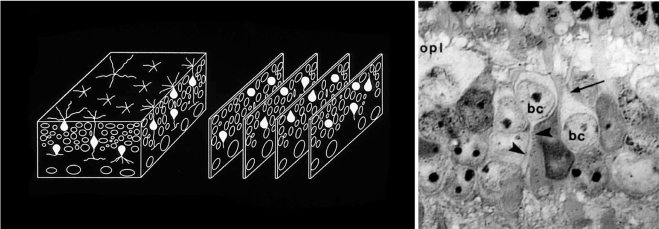
Defining the major classes of interneurons. Horizontal, Müller, bipolar, and amacrine cells are not readily distinguishable in ordinary histologic stains. To inventory the specific types of retinal neurons, it was first necessary to establish a baseline—to quantitatively describe the major classes of retinal interneurons. We devised a way to count the cells independent of any staining methodology. The inner plexiform layer was treated as a three-dimensional solid. Samples (blocks of tissue) from locations around the retina were serially sectioned, and every cell within each block was identified. The identification was made by visualizing their major processes at their exit from the cell bodies (right). Contrary to earlier belief, the retinas of different mammalian species had very similar cell populations, as shown in Figure 3. This quantitative inventory of the major cell classes could be used as a foundation on which the inventories of individual bipolar and amacrine cell populations could be built. OPL, outer plexiform layer; BC, bipolar cell; arrow, bipolar cell dendrite; arrowhead, bipolar cell axon. Left: Adapted from Masland RH. Neuronal diversity in the retina. Current Opin Neurobiol. 2001;11:431–436. © Elsevier. Right: Adapted from Strettoi E, Masland RH. The organization of the inner nuclear layer of the rabbit retina. J Neurosci. 1995;15:875–888. ©Society for Neuroscience.
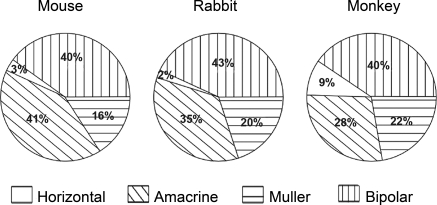
The populations of retinal interneurons in the mouse, rabbit, and monkey. Data for the mouse and the rabbit were obtained using the serial reconstruction method shown in Figure 2. Data from the monkey are from Martin and Grünert. The comparison is made for regions of retina in the three species where the density of cones is similar—from the central retina in the rabbit and mouse and the midperiphery in the monkey. Horizontal cells in monkeys are somewhat more numerous (they are also smaller in lateral extent) than are horizontal cells in mice and rabbits. What is most striking about the comparison among the three species, however, is their overall similarity.

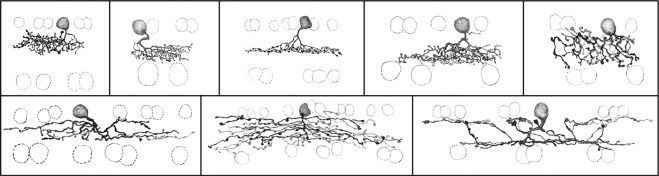
The types of bipolar cells in the rabbit retina. A more recent study in the mouse retina, using XFP-expressing transgenic animal strains and immunostaining to distinguish the bipolar cell types, also arrived at 12 types of cone bipolar cells. These are drawings of individual Golgi-stained cells. They are representative of a much larger sample. The 12 types of cone bipolars illustrated here (and one rod bipolar) were classified by study of a large population of cells stained by the photofilling method, by Golgi staining, and by injection with Lucifer yellow after recording. Most have narrow dendritic and axonal fields, but one (wide-field Cba2) contacts the sparse blue cones exclusively and has correspondingly wide arbors. The naming of the cells is not definitive; it is likely that the final nomenclature will use either the names of the cells' molecular markers or a simple numbering.
Some of the types of amacrine cell present in the rabbit retina. This is a subset of the total population of amacrine cells distinguishable in photofilling and Golgi stains. In the initial study, 29 types of amacrine cell were identified., Later work indicated that the number of types of wide-field amacrine cells had been underestimated, because a small number of wide-field cells of any one type can still blanket the retinal surface completely, and they are thus rarely encountered by any sampling method. Note that many of the amacrine cells shown here violate the division of the inner plexiform layer into ON (inner) and OFF (outer) halves: They carry ON information to the OFF cells, and OFF information to the ON cells.
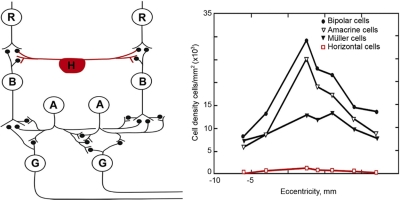
Despite their prominent place in the textbook representation of the retina, horizontal cells are a numerically insignificant fraction of the retina's total interneurons. Left: this classic drawing (courtesy of Dr. John Dowling) accurately conveys the main types of retinal neuron and their synaptic interrelations. What this type of drawing cannot show is the relative number of the cells, which adds an additional dimension to our understanding. The counts are shown at right, from serial reconstructions as shown in Figure 2 and 3. The lateral connections of the retina are dominated quantitatively by amacrine cells. R, photoreceptor cell; H, horizontal cell; B, bipolar cell; A, amacrine cell; G, ganglion cell.
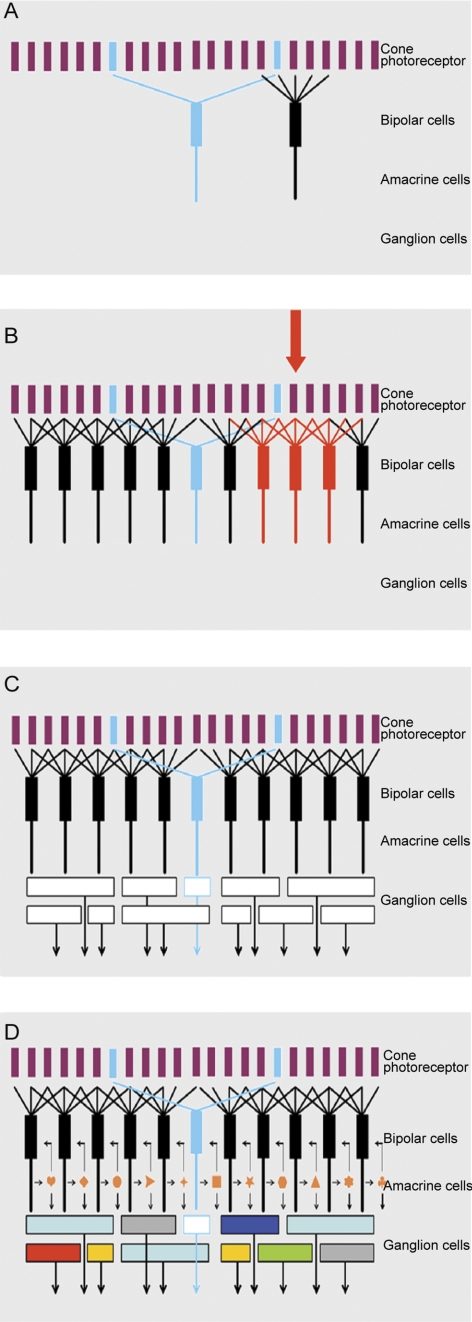
The four panels of this figure show exploded views of the retina's circuitry. Here, the mosaic of cone photoreceptors is shown at the top. (The late-evolving rod system is not shown, as it is a minor part of the retina's circuitry, and is used only in deep darkness.) The mosaic of cone photoreceptors is sampled by the “diffuse” bipolar cells (black), which receive input from all the cones within their reach, and by a small group of “blue bipolars,” which selectively contact the infrequent blue cones. This selectivity preserves the chromatic purity of the responses of the blue bipolar cells. Later retinal circuitry (not shown) compares the output of the blue (short wavelength) cones to that of the long wavelength cones. This comparison is the fundamental basis of mammalian color vision. Figure 7B. The diffuse bipolar cells are far more numerous than the blue bipolars and come in 11 functional types, each transmitting a different component of the cone's output. Stimulation of an individual cone (red arrow) transmits information to all the bipolar cells that contact it (red bipolar cells in the illustration). In this case the red bipolar cells would all be of one functional type; for example, all of them would be ON-transient bipolars. But that same (arrow) cone would also be contacted by bipolar cells of the other 10 types, carrying different components of the cone's output. The whole array cannot be shown in a planar diagram like this one—there is not enough space for all the bipolar cells in a single plane, and 12 different colors of bipolar cell would be required. Figure 7C. The different types of bipolar cells synapse on different types of ganglion cells. Since each type of bipolar cell conveys a different type of information to the inner retina, each conveys its own particular version of the visual input to the ganglion cell(s) on which it synapses. This is the initial step in the creation of ganglion cell diversity. In the example, the blue bipolar cell synapses on a specific type of ganglion cell, which then becomes a blue-sensitive ganglion cell. The same is true for each of the other bipolar cell types. If this were the whole story, the organization of the inner retina would be relatively simple. There might still be 12 (or more) functional types of ganglion cells, but the bipolar cells would provide the only drive to the ganglion cells, so that the ganglion cell response would be determined purely by the response tuning of the bipolar cell, the physiology of the bipolar cell synapse and the ganglion cell's ion channels. Figure 7D. The final responses of the ganglion cells are highly selective: Some respond only to a particular direction of stimulus movement, some specifically to low or to high contrast, some specifically to rapid movement, and so on. This final diversity is created by the action of amacrine cells. Amacrine cells receive a direct excitatory input from bipolar cells, and they make inhibitory synaptic inputs back onto the axon terminals of the bipolar cells. This is a classic feedback system, creating a control point at the bottleneck where information enters the inner retina from the outer retina. However, amacrine cells also synapse on each other; and they feed forward on retinal ganglion cells. These feed-forward synapses are responsible for some of the more interesting properties of the ganglion cell response. A famous case is the starburst amacrine cell, which creates direction selectivity in one type of retinal ganglion cell., Not shown are the wide-field amacrine cells, which can span the whole width of the retinal surface.
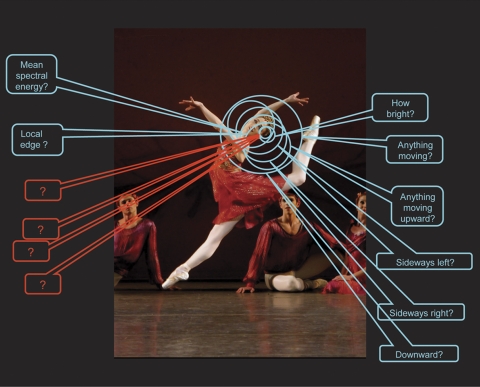
The retina interrogates any point in visual space about a number of distinct qualities. Retinal ganglion cells tile the retinal surface: Each of the ∼20 distinct types of retinal ganglion cell covers the retinal surface uniformly, and this mosaic is independent of the mosaics of the other 19 types. Thus, at every point on the retina at least one example of each of the 20 ganglion cell types is present. Correspondingly, a point in visual space is surveyed by at least one of each of the types of retinal ganglion cell, each reporting to the brain a different feature of the visual world. In this example, we consider a small region on the trailing side of the dancer's head. Different retinal ganglion cells report on whether there is, within that area of space, movement, movement in a particular direction, an object of a particular spectral composition, among other reportings. The nature of eight of these reportings are known (there are eight well-characterized types of ganglion cell in the rabbit retina, on which the illustration is based). Many more are known to exist, from the presence of structurally distinct ganglion cell types, but their physiology remains to be characterized. From structural evidence, the number of encodings appears to be similar in all mammalian retinas. Image: New York City Ballet's Ashley Bouder in Jerome Robbins' “The Four Seasons.” Photo credit: Paul Kolnik.
References
-
- Cajal SR. The Structure of the Retina. 1892 Translated by Thorpe SA, Glickstein M. Springfield, IL: Thomas; 1972
-
- Boycott BB, Dowling JE. Organization of the primate retina: light microscopy. Phil Trans R Soc Lond Biol B. 1969;255:109–184 - PubMed
-
- Kolb H, Nelson R, Mariani A. Amacrine cells, bipolar cells and ganglion cells of the cat retina: a Golgi study. Vision Res. 1981;21:1081–1114 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
