

Diverse two-cysteine photocycles in phytochromes and cyanobacteriochromes
- PMID: 21712441
- PMCID: PMC3141974
- DOI: 10.1073/pnas.1107844108
Diverse two-cysteine photocycles in phytochromes and cyanobacteriochromes
Abstract
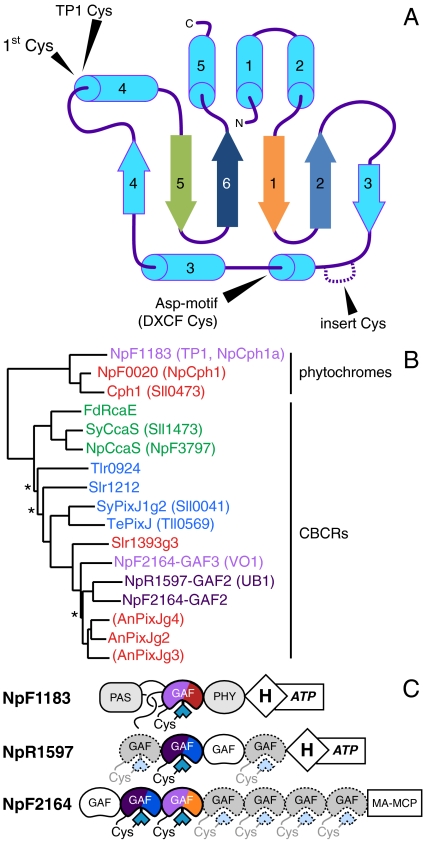
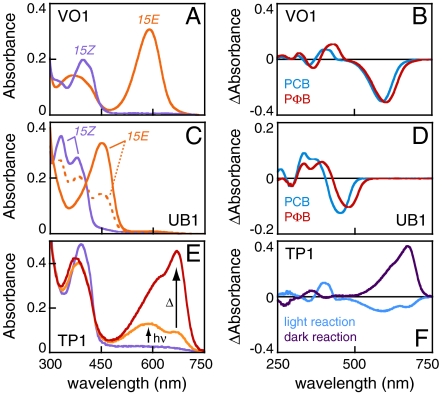
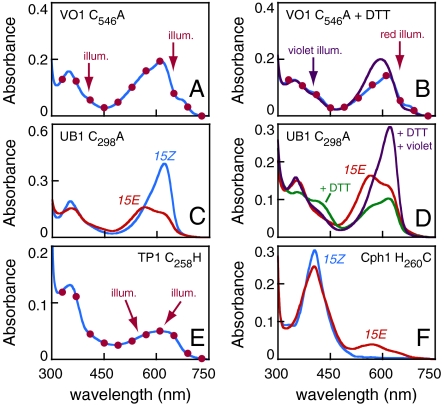
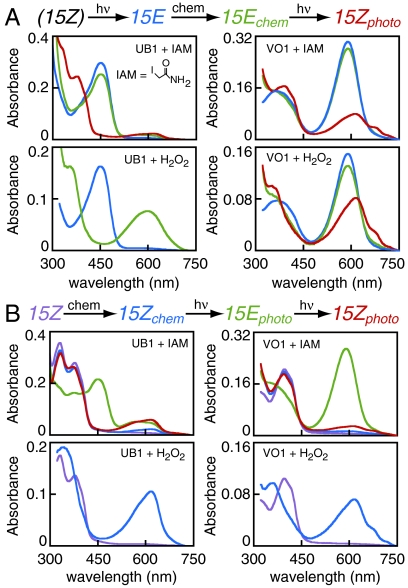
Phytochromes are well-known as photoactive red- and near IR-absorbing chromoproteins with cysteine-linked linear tetrapyrrole (bilin) prosthetic groups. Phytochrome photoswitching regulates adaptive responses to light in both photosynthetic and nonphotosynthetic organisms. Exclusively found in cyanobacteria, the related cyanobacteriochrome (CBCR) sensors extend the photosensory range of the phytochrome superfamily to shorter wavelengths of visible light. Blue/green light sensing by a well-studied subfamily of CBCRs proceeds via a photolabile thioether linkage to a second cysteine fully conserved in this subfamily. In the present study, we show that dual-cysteine photosensors have repeatedly evolved in cyanobacteria via insertion of a second cysteine at different positions within the bilin-binding GAF domain (cGMP-specific phosphodiesterases, cyanobacterial adenylate cyclases, and formate hydrogen lyase transcription activator FhlA) shared by CBCRs and phytochromes. Such sensors exhibit a diverse range of photocycles, yet all share ground-state absorbance of near-UV to blue light and a common mechanism of light perception: reversible photoisomerization of the bilin 15,16 double bond. Using site-directed mutagenesis, chemical modification and spectroscopy to characterize novel dual-cysteine photosensors from the cyanobacterium Nostoc punctiforme ATCC 29133, we establish that this spectral diversity can be tuned by varying the light-dependent stability of the second thioether linkage. We also show that such behavior can be engineered into the conventional phytochrome Cph1 from Synechocystis sp. PCC6803. Dual-cysteine photosensors thus allow the phytochrome superfamily in cyanobacteria to sense the full solar spectrum at the earth surface from near infrared to near ultraviolet.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Montgomery BL, Lagarias JC. Phytochrome ancestry. Sensors of bilins and light. Trends Plant Sci. 2002;7:357–366. - PubMed
-
- Auldridge ME, Forest KT. Bacterial phytochromes: More than meets the light. Crit Rev Biochem Mol Biol. 2011;46:67–88. - PubMed
-
- Giraud E, et al. Bacteriophytochrome controls photosystem synthesis in anoxygenic bacteria. Nature. 2002;417:202–205. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
