

Over-expression of mitochondrial ferritin affects the JAK2/STAT5 pathway in K562 cells and causes mitochondrial iron accumulation
- PMID: 21712541
- PMCID: PMC3186302
- DOI: 10.3324/haematol.2011.042952
Over-expression of mitochondrial ferritin affects the JAK2/STAT5 pathway in K562 cells and causes mitochondrial iron accumulation
Abstract
Background: Mitochondrial ferritin is a nuclear encoded iron-storage protein localized in mitochondria. It has anti-oxidant properties related to its ferroxidase activity, and it is able to sequester iron avidly into the organelle. The protein has a tissue-specific pattern of expression and is also highly expressed in sideroblasts of patients affected by hereditary sideroblastic anemia and by refractory anemia with ringed sideroblasts. The present study examined whether mitochondrial ferritin has a role in the pathogenesis of these diseases.
Design and methods: We analyzed the effect of mitochondrial ferritin over-expression on the JAK2/STAT5 pathway, on iron metabolism and on heme synthesis in erythroleukemic cell lines. Furthermore its effect on apoptosis was evaluated on human erythroid progenitors.
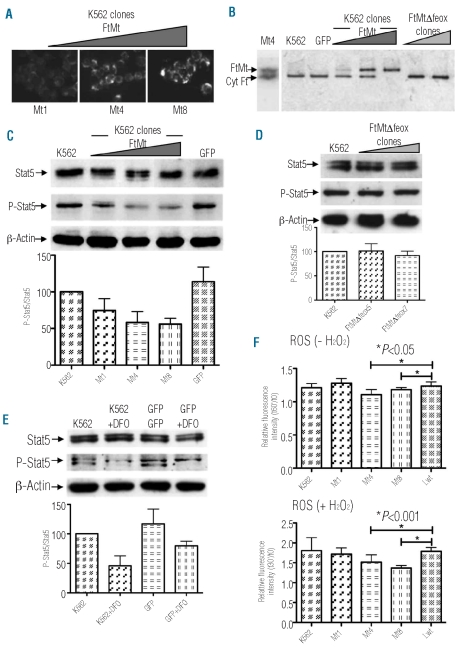
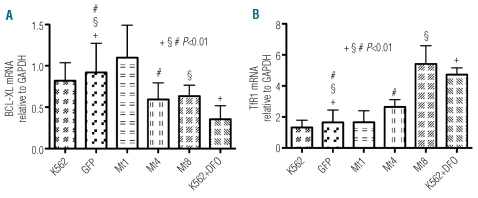
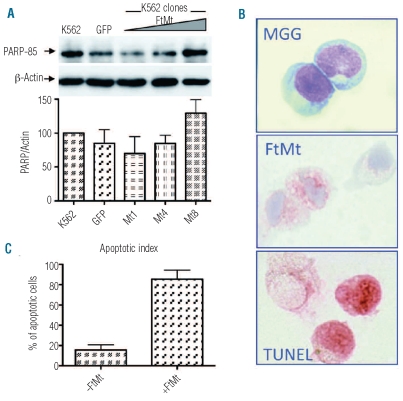
Results: Data revealed that a high level of mitochondrial ferritin reduced reactive oxygen species and Stat5 phosphorylation while promoting mitochondrial iron loading and cytosolic iron starvation. The decline of Stat5 phosphorylation induced a decrease of the level of anti-apoptotic Bcl-xL transcript compared to that in control cells; however, transferrin receptor 1 transcript increased due to the activation of the iron responsive element/iron regulatory protein machinery. Also, high expression of mitochondrial ferritin increased apoptosis, limited heme synthesis and promoted the formation of Perls-positive granules, identified by electron microscopy as iron granules in mitochondria.
Conclusions: Our results provide evidence suggesting that Stat5-dependent transcriptional regulation is displaced by strong cytosolic iron starvation status induced by mitochondrial ferritin. The protein interferes with JAK2/STAT5 pathways and with the mechanism of mitochondrial iron accumulation.
Figures

References
-
- Levi S, Corsi B, Bosisio M, Invernizzi R, Volz A, Sanford D, et al. A human mitochondrial ferritin encoded by an intronless gene. J Biol Chem. 2001;276(27):24437–40. - PubMed
-
- Arosio P, Levi S. Cytosolic and mitochondrial ferritins in the regulation of cellular iron homeostasis and oxidative damage. Biochim Biophys Acta. 2010;1800(8):783–92. - PubMed
-
- Drysdale J, Arosio P, Invernizzi R, Cazzola M, Volz A, Corsi B, et al. Mitochondrial ferritin: a new player in iron metabolism. Blood Cells Mol Dis. 2002;29(3):376–83. - PubMed
-
- Langlois d'Estaintot B, Santambrogio P, Granier T, Gallois B, Chevalier JM, Precigoux G, et al. Crystal structure and biochemical properties of the human mitochondrial ferritin and its mutant Ser144Ala. J Mol Biol. 2004;340(2):277–93. - PubMed
-
- Bou-Abdallah F, Santambrogio P, Levi S, Arosio P, Chasteen ND. Unique iron binding and oxidation properties of human mitochondrial ferritin: a comparative analysis with human H-chain ferritin. J Mol Biol. 2005;347(3):543–54. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
