

RNA folding pathways and the self-assembly of ribosomes
- PMID: 21714483
- PMCID: PMC4361232
- DOI: 10.1021/ar2000474
RNA folding pathways and the self-assembly of ribosomes
Abstract
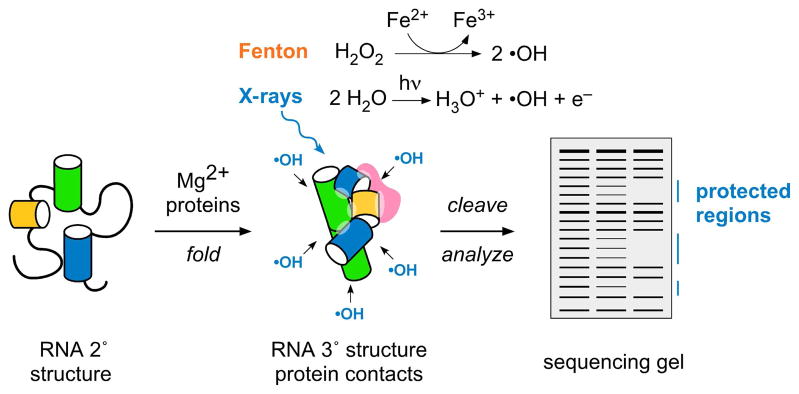
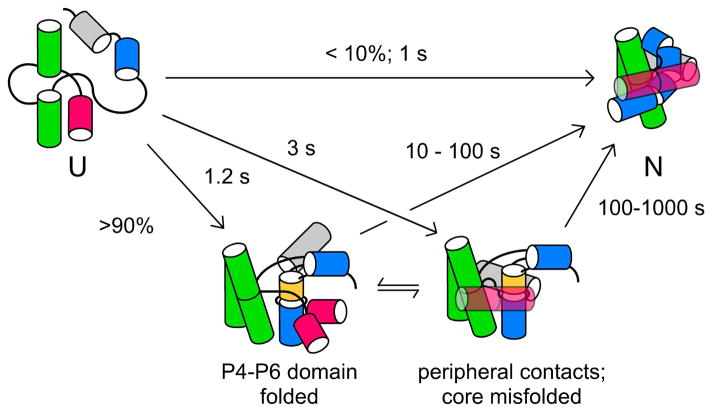
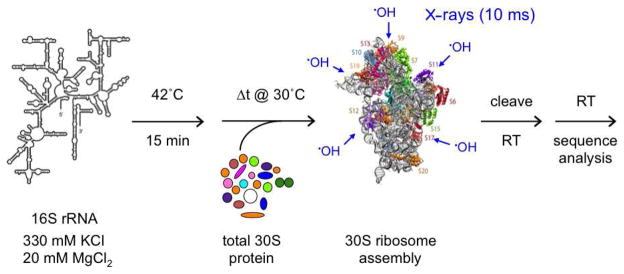
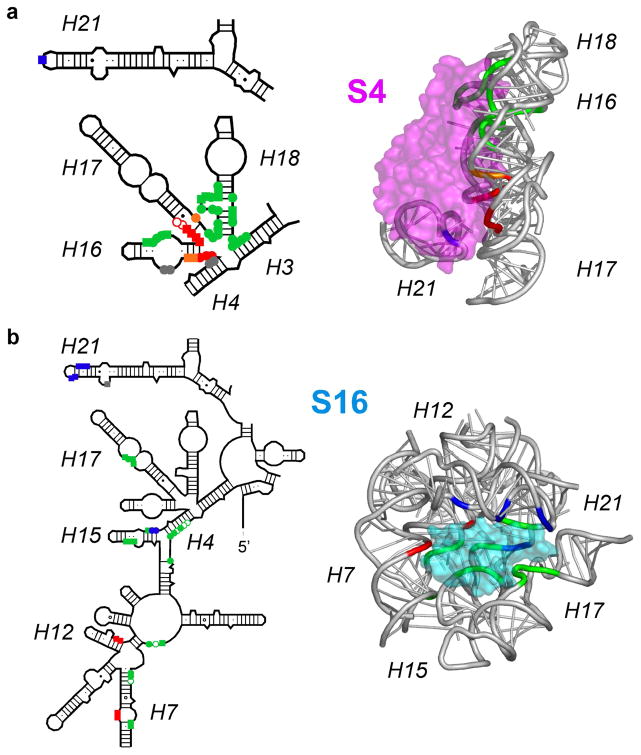
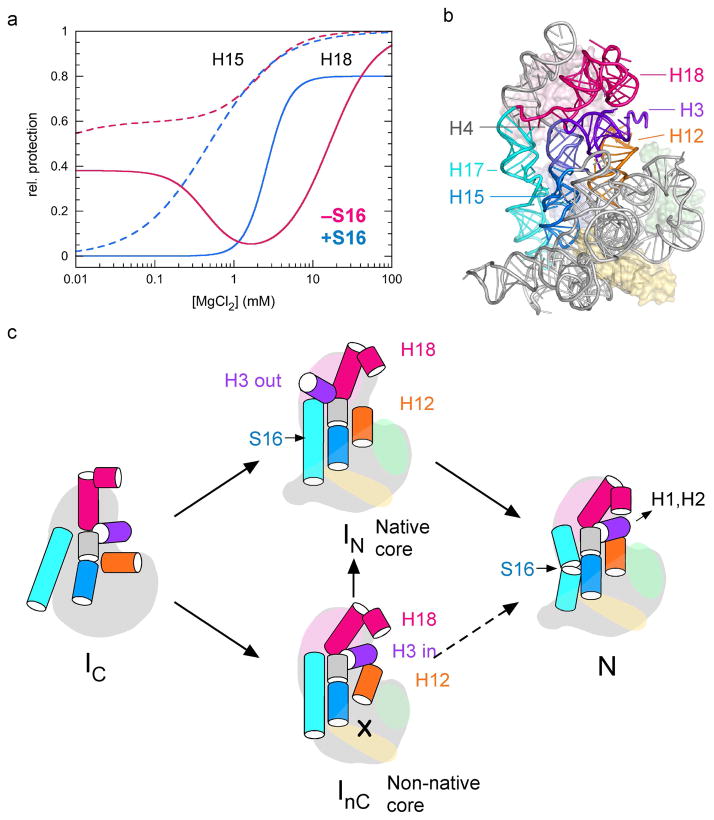
Many RNAs do not directly code proteins but are nonetheless indispensable to cellular function. These strands fold into intricate three-dimensional shapes that are essential structures in protein synthesis, splicing, and many other processes of gene regulation and expression. A variety of biophysical and biochemical methods are now showing, in real time, how ribosomal subunits and other ribonucleoprotein complexes assemble from their molecular components. Footprinting methods are particularly useful for studying the folding of long RNAs: they provide quantitative information about the conformational state of each residue and require little material. Data from footprinting complement the global information available from small-angle X-ray scattering or cryo-electron microscopy, as well as the dynamic information derived from single-molecule Förster resonance energy transfer (FRET) and NMR methods. In this Account, I discuss how we have used hydroxyl radical footprinting and other experimental methods to study pathways of RNA folding and 30S ribosome assembly. Hydroxyl radical footprinting probes the solvent accessibility of the RNA backbone at each residue in as little as 10 ms, providing detailed views of RNA folding pathways in real time. In conjunction with other methods such as solution scattering and single-molecule FRET, time-resolved footprinting of ribozymes showed that stable domains of RNA tertiary structure fold in less than 1 s. However, the free energy landscapes for RNA folding are rugged, and individual molecules kinetically partition into folding pathways that lead through metastable intermediates, stalling the folding or assembly process. Time-resolved footprinting was used to follow the formation of tertiary structure and protein interactions in the 16S ribosomal RNA (rRNA) during the assembly of 30S ribosomes. As previously observed in much simpler ribozymes, assembly occurs in stages, with individual molecules taking different routes to the final complex. Interactions occur concurrently in all domains of the 16S rRNA, and multistage protection of binding sites of individual proteins suggests that initial encounter complexes between the rRNA and ribosomal proteins are remodeled during assembly. Equilibrium footprinting experiments showed that one primary binding protein was sufficient to stabilize the tertiary structure of the entire 16S 5'-domain. The rich detail available from the footprinting data showed that the secondary assembly protein S16 suppresses non-native structures in the 16S 5'-domain. In doing so, S16 enables a conformational switch distant from its own binding site, which may play a role in establishing interactions with other domains of the 30S subunit. Together, the footprinting results show how protein-induced changes in RNA structure are communicated over long distances, ensuring cooperative assembly of even very large RNA-protein complexes such as the ribosome.
Figures
Similar articles
-
Concurrent nucleation of 16S folding and induced fit in 30S ribosome assembly.Nature. 2008 Oct 30;455(7217):1268-72. doi: 10.1038/nature07298. Epub 2008 Sep 10. Nature. 2008. PMID: 18784650 Free PMC article.
-
S16 throws a conformational switch during assembly of 30S 5' domain.Nat Struct Mol Biol. 2009 Apr;16(4):438-45. doi: 10.1038/nsmb.1585. Epub 2009 Apr 3. Nat Struct Mol Biol. 2009. PMID: 19343072 Free PMC article.
-
A metastable rRNA junction essential for bacterial 30S biogenesis.Nucleic Acids Res. 2018 Jun 1;46(10):5182-5194. doi: 10.1093/nar/gky120. Nucleic Acids Res. 2018. PMID: 29850893 Free PMC article.
-
RNA folding and ribosome assembly.Curr Opin Chem Biol. 2008 Dec;12(6):667-73. doi: 10.1016/j.cbpa.2008.09.024. Epub 2008 Oct 18. Curr Opin Chem Biol. 2008. PMID: 18935976 Free PMC article. Review.
-
RNA-protein interactions in 30S ribosomal subunits: folding and function of 16S rRNA.Science. 1989 May 19;244(4906):783-90. doi: 10.1126/science.2658053. Science. 1989. PMID: 2658053 Review.
Cited by
-
Cryo-EM structures of the late-stage assembly intermediates of the bacterial 50S ribosomal subunit.Nucleic Acids Res. 2013 Aug;41(14):7073-83. doi: 10.1093/nar/gkt423. Epub 2013 May 21. Nucleic Acids Res. 2013. PMID: 23700310 Free PMC article.
-
Transcription complexes as RNA chaperones.Transcription. 2021 Aug;12(4):126-155. doi: 10.1080/21541264.2021.1985931. Epub 2021 Nov 1. Transcription. 2021. PMID: 34719334 Free PMC article.
-
Nanopore electric snapshots of an RNA tertiary folding pathway.Nat Commun. 2017 Nov 13;8(1):1458. doi: 10.1038/s41467-017-01588-z. Nat Commun. 2017. PMID: 29133841 Free PMC article.
-
Evidence that viral RNAs have evolved for efficient, two-stage packaging.Proc Natl Acad Sci U S A. 2012 Sep 25;109(39):15769-74. doi: 10.1073/pnas.1204357109. Epub 2012 Sep 10. Proc Natl Acad Sci U S A. 2012. PMID: 23019360 Free PMC article.
-
Native Purification and Analysis of Long RNAs.Methods Enzymol. 2015;558:3-37. doi: 10.1016/bs.mie.2015.01.008. Epub 2015 Feb 27. Methods Enzymol. 2015. PMID: 26068736 Free PMC article.
References
-
- Serganov A. Determination of riboswitch structures: light at the end of the tunnel? RNA Biol. 2010;7:98–103. - PubMed
-
- Ramakrishnan V. Ribosome structure and the mechanism of translation. Cell. 2002;108:557–72. - PubMed
-
- Fedor MJ. Comparative enzymology and structural biology of RNA self-cleavage. Annu Rev Biophys. 2009;38:271–99. - PubMed
-
- Newman AJ, Nagai K. Structural studies of the spliceosome: blind men and an elephant. Curr Opin Struct Biol. 2010;20:82–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
