

Mechanism of neutralization by the broadly neutralizing HIV-1 monoclonal antibody VRC01
- PMID: 21715490
- PMCID: PMC3165784
- DOI: 10.1128/JVI.00754-11
Mechanism of neutralization by the broadly neutralizing HIV-1 monoclonal antibody VRC01
Abstract
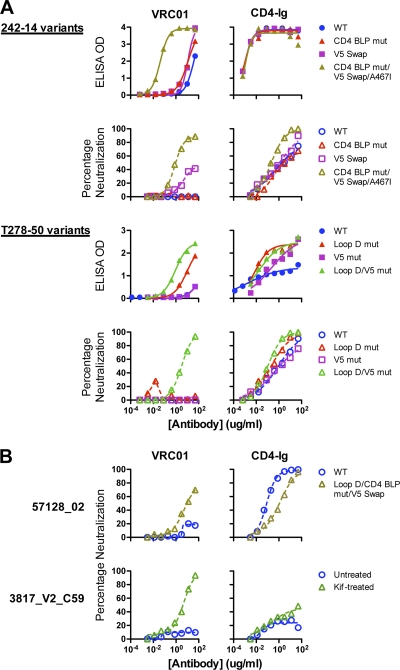
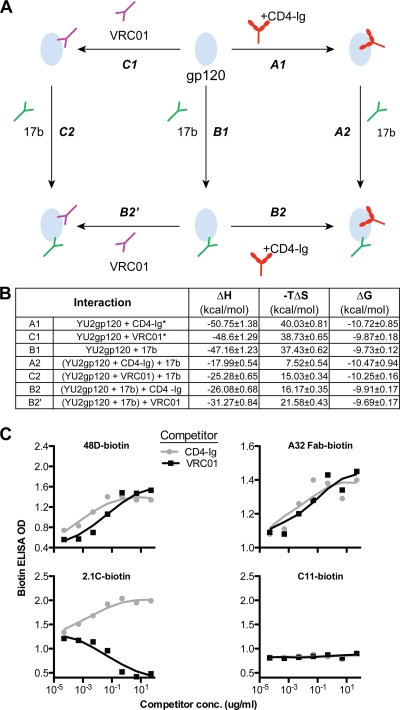
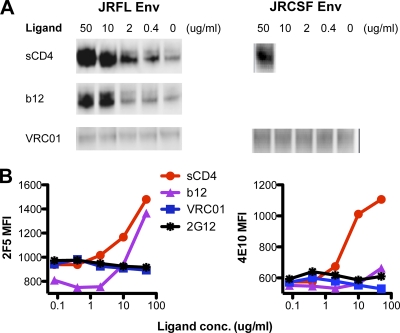
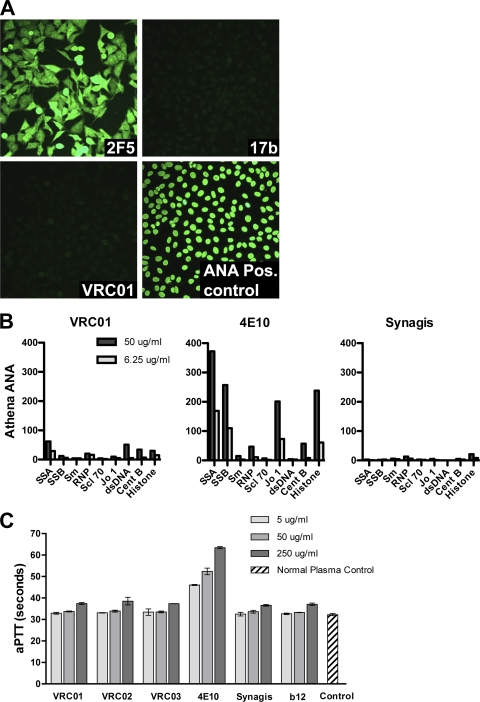
The structure of VRC01 in complex with the HIV-1 gp120 core reveals that this broadly neutralizing CD4 binding site (CD4bs) antibody partially mimics the interaction of the primary virus receptor, CD4, with gp120. Here, we extended the investigation of the VRC01-gp120 core interaction to the biologically relevant viral spike to better understand the mechanism of VRC01-mediated neutralization and to define viral elements associated with neutralization resistance. In contrast to the interaction of CD4 or the CD4bs monoclonal antibody (MAb) b12 with the HIV-1 envelope glycoprotein (Env), occlusion of the VRC01 epitope by quaternary constraints was not a major factor limiting neutralization. Mutagenesis studies indicated that VRC01 contacts within the gp120 loop D, the CD4 binding loop, and the V5 region were necessary for optimal VRC01 neutralization, as suggested by the crystal structure. In contrast to interactions with the soluble gp120 monomer, VRC01 interaction with the native viral spike did not occur in a CD4-like manner; VRC01 did not induce gp120 shedding from the Env spike or enhance gp41 membrane proximal external region (MPER)-directed antibody binding to the Env spike. Finally, VRC01 did not display significant reactivity with human antigens, boding well for potential in vivo applications. The data indicate that VRC01 interacts with gp120 in the context of the functional spike in a manner distinct from that of CD4. It achieves potent neutralization by precisely targeting the CD4bs without requiring alterations of Env spike configuration and by avoiding steric constraints imposed by the quaternary structure of the functional Env spike.
Figures
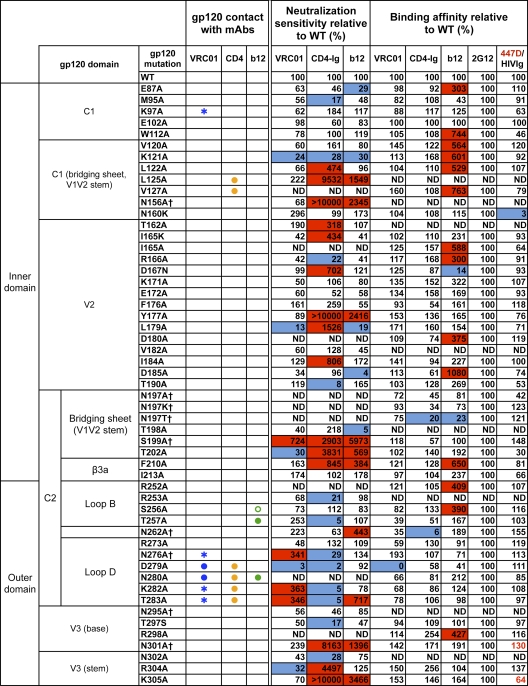
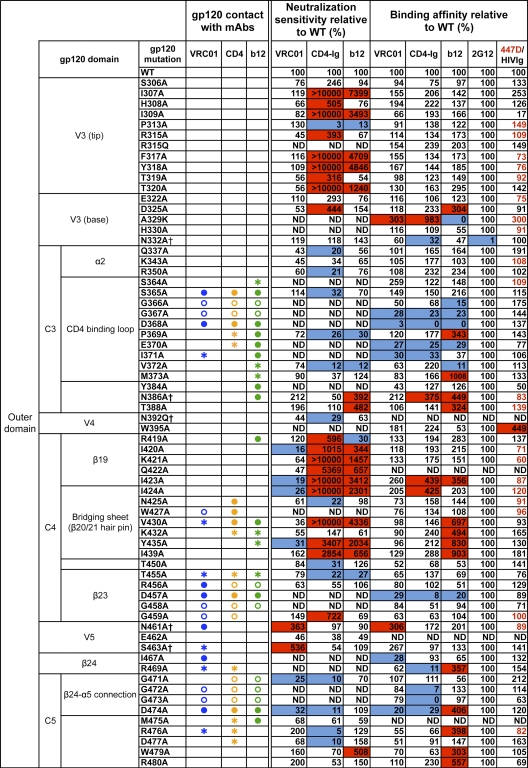
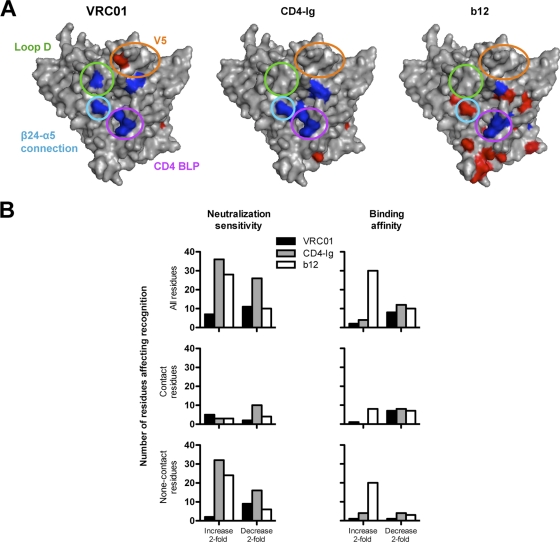
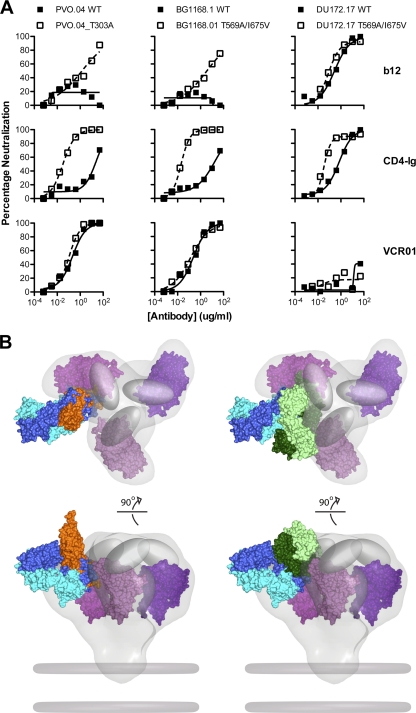
References
-
- Alkhatib G., et al. 1996. CC CKR5: a RANTES, MIP-1alpha, MIP-1beta receptor as a fusion cofactor for macrophage-tropic HIV-1. Science 272:1955–1958 - PubMed
-
- Buchacher A., et al. 1994. Generation of human monoclonal antibodies against HIV-1 proteins; electrofusion and Epstein-Barr virus transformation for peripheral blood lymphocyte immortalization. AIDS Res. Hum. Retroviruses 10:359–369 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
