

17(R)-resolvin D1 specifically inhibits transient receptor potential ion channel vanilloid 3 leading to peripheral antinociception
- PMID: 21718307
- PMCID: PMC3315040
- DOI: 10.1111/j.1476-5381.2011.01568.x
17(R)-resolvin D1 specifically inhibits transient receptor potential ion channel vanilloid 3 leading to peripheral antinociception
Abstract
Background and purpose: Transient receptor potential ion channel vanilloid 3 (TRPV3) is expressed in skin keratinocytes and plays an important role in thermal and chemical nociceptions in the periphery. The presence of TRPV3 inhibitors would improve our understanding of TRPV3 function and help to develop receptor-specific analgesics. However, little is known about physiological substances that specifically inhibit TRPV3 activity. Here, we investigated whether 17(R)-resolvin D1 (17R-RvD1), a naturally occurring pro-resolving lipid specifically affects TRPV3 activity.
Experimental approach: We examined the effect of 17R-RvD1 on sensory TRP channels using Ca(2+) imaging and whole cell electrophysiology experiments in a HEK cell heterologous expression system, cultured sensory neurons and keratinocytes. We also examined changes in sensory TRP agonist-specific acute licking/flicking or flinching behaviours and mechanical and thermal pain behaviours using Hargreaves, Randall-Selitto and von Frey assay systems in the absence and presence of inflammation.
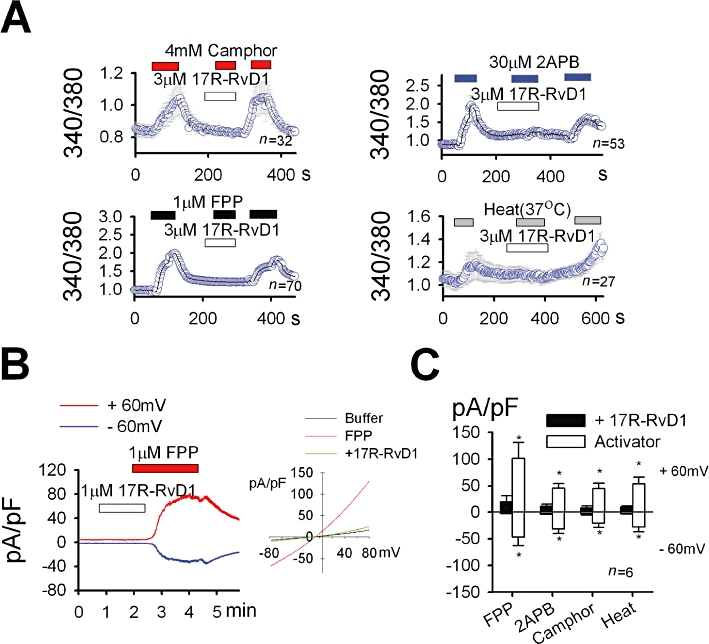
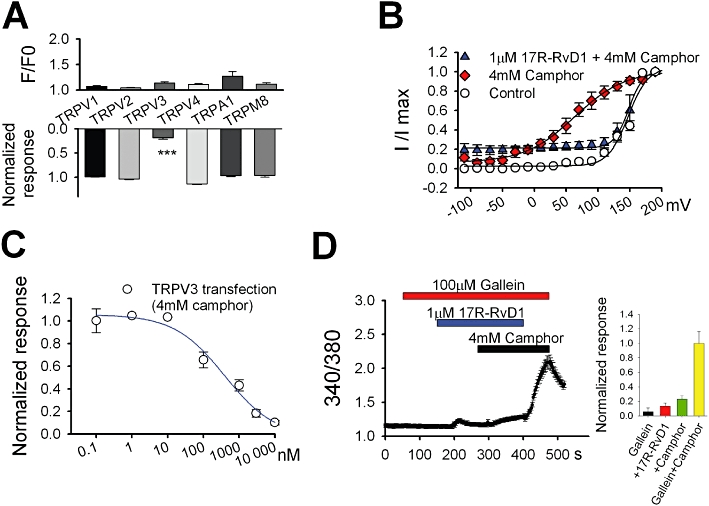
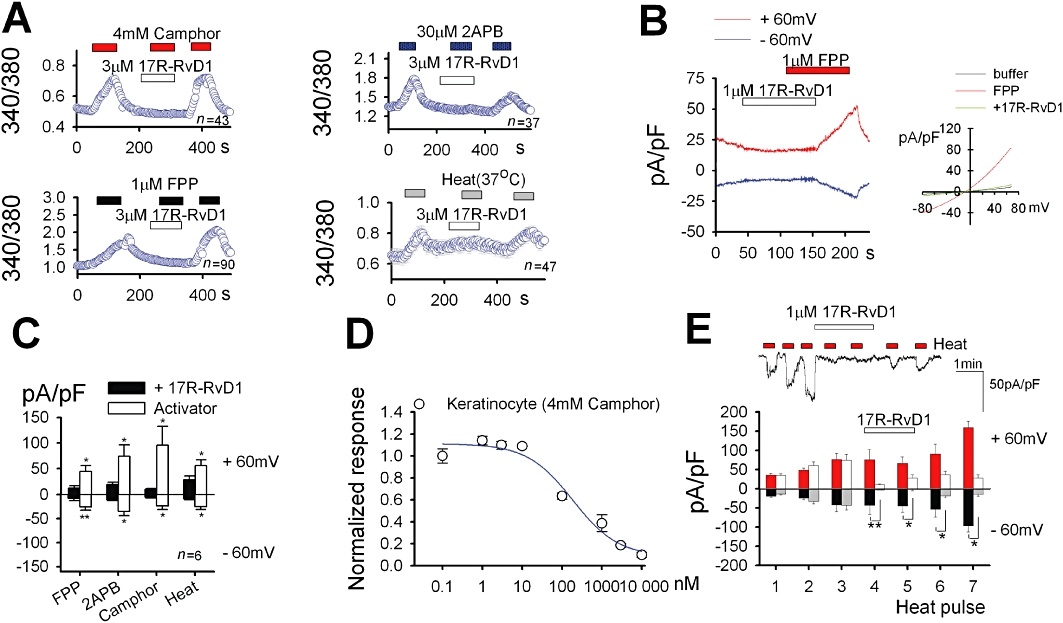
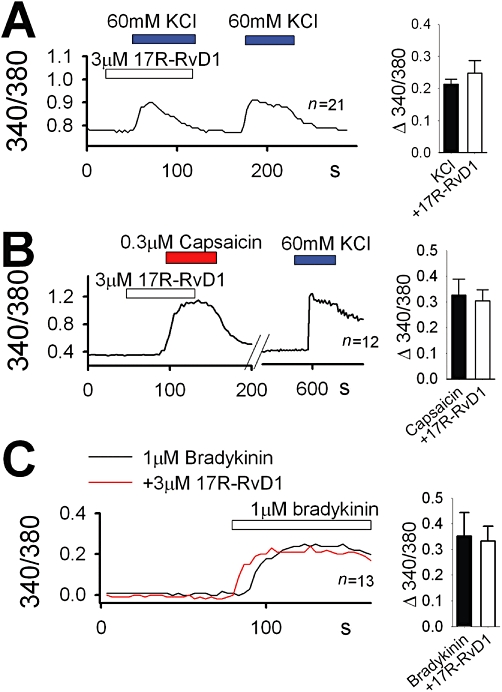
Key results: We showed that 17R-RvD1 specifically suppresses TRPV3-mediated activity at nanomolar and micromolar concentrations. The voltage-dependence of TRPV3 activation by camphor was shifted rightwards by 17R-RvD1, which indicates its inhibitory mechanism is as a result of a shift in voltage-dependence. Consistently, TRPV3-specific acute pain behaviours were attenuated by locally injected 17R-RvD1. Moreover, the administration of 17R-RvD1 significantly reversed the thermal hypersensitivity that occurs during an inflammatory response. Knockdown of epidermal TRPV3 blunted these antinociceptive effects of 17R-RvD1.
Conclusions and implications: 17R-RvD1 is a novel natural inhibitory substance specific for TRPV3. The results of our behavioural studies suggest that 17R-RvD1 has acute analgesic potential via TRPV3-specific mechanisms.
© 2011 The Authors. British Journal of Pharmacology © 2011 The British Pharmacological Society.
Figures
References
-
- Ariel A, Serhan CN. Resolvins and protectins in the termination program of acute inflammation. Trends Immunol. 2007;28:176–183. - PubMed
-
- Bandell M, Story GM, Hwang SW, Viswanath V, Eid SR, Petrus MJ, et al. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron. 2004;41:849–857. - PubMed
-
- Bang S, Kim KY, Yoo S, Kim YG, Hwang SW. Transient receptor potential A1 mediates acetaldehyde-evoked pain sensation. Eur J Neurosci. 2007a;26:2516–2523. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
