

Gonadotropin-regulated testicular RNA helicase (GRTH/DDX25), a negative regulator of luteinizing/chorionic gonadotropin hormone-induced steroidogenesis in Leydig cells: central role of steroidogenic acute regulatory protein (StAR)
- PMID: 21719703
- PMCID: PMC3191034
- DOI: 10.1074/jbc.M111.236083
Gonadotropin-regulated testicular RNA helicase (GRTH/DDX25), a negative regulator of luteinizing/chorionic gonadotropin hormone-induced steroidogenesis in Leydig cells: central role of steroidogenic acute regulatory protein (StAR)
Abstract
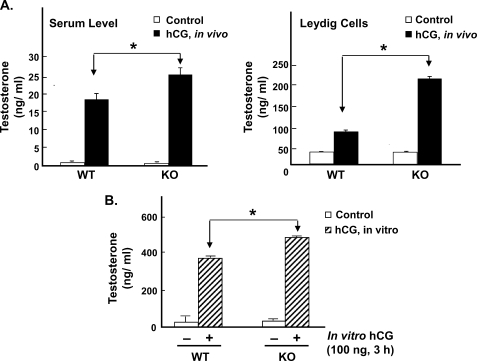
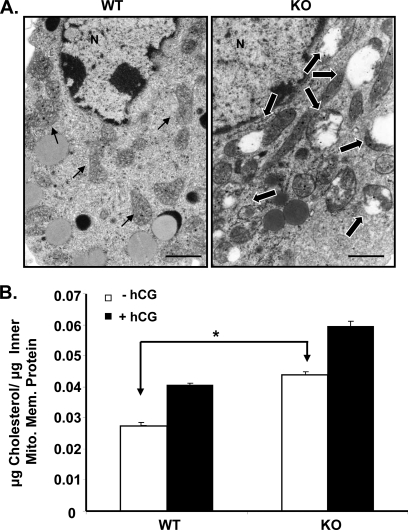
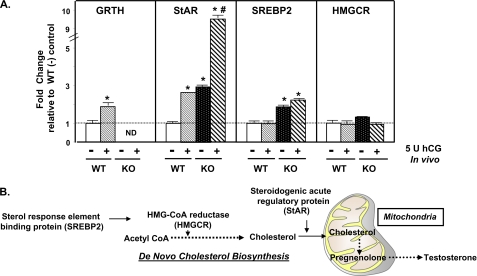
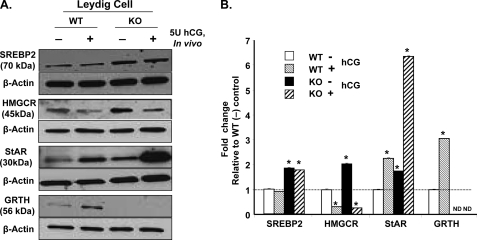
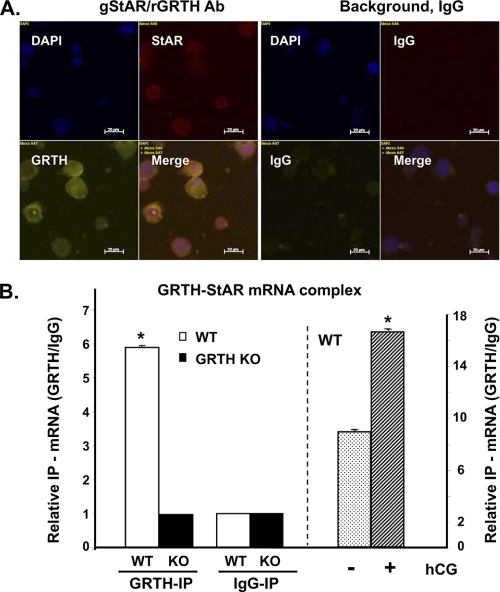
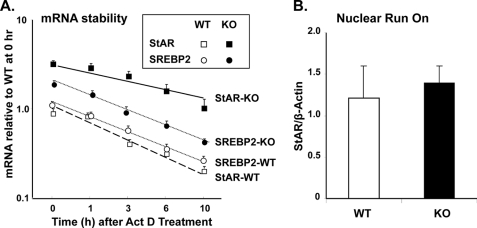
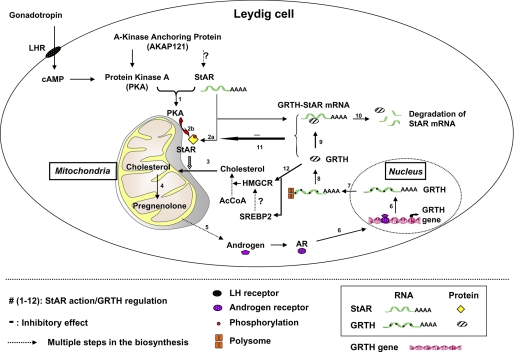
Gonadotropin-regulated testicular RNA helicase (GRTH/DDX25) is a testis-specific gonadotropin-regulated RNA helicase that is present in Leydig cells (LCs) and germ cells and is essential for spermatid development and completion of spermatogenesis. Normal basal levels of testosterone in serum and LCs were observed in GRTH null (GRTH(-/-)) mice. However, testosterone production was enhanced in LCs of GRTH(-/-) mice compared with WT mice by both in vivo and in vitro human chorionic gonadotropin stimulation. LCs of GRTH(-/-) mice had swollen mitochondria with a significantly increased cholesterol content in the inner mitochondrial membrane. Basal protein levels of SREBP2, HMG-CoA reductase, and steroidogenic acute regulatory protein (StAR; a protein that transports cholesterol to the inner mitochondrial membrane) were markedly increased in LCs of GRTH(-/-) mice compared with WT mice. Gonadotropin stimulation caused an increase in StAR mRNA levels and protein expression in GRTH(-/-) mice versus WT mice, with no further increase in SREBP2 and down-regulation of HMG-CoA reductase protein. The half-life of StAR mRNA was significantly increased in GRTH(-/-) mice. Moreover, association of StAR mRNA with GRTH protein was observed in WT mice. Human chorionic gonadotropin increased GRTH gene expression and its associated StAR protein at cytoplasmic sites. Taken together, these findings indicate that, through its negative role in StAR message stability, GRTH regulates cholesterol availability at the mitochondrial level. The finding of an inhibitory action of GRTH associated with gonadotropin-mediated steroidogenesis has provided insights into a novel negative autocrine molecular control mechanism of this helicase in the regulation of steroid production in the male.
Figures
Similar articles
-
Role of gonadotropin regulated testicular RNA helicase (GRTH/Ddx25) on polysomal associated mRNAs in mouse testis.PLoS One. 2012;7(3):e32470. doi: 10.1371/journal.pone.0032470. Epub 2012 Mar 30. PLoS One. 2012. PMID: 22479328 Free PMC article.
-
A 5'-flanking region of gonadotropin-regulated testicular RNA helicase (GRTH/DDX25) gene directs its cell-specific androgen-regulated gene expression in testicular germ cells.Endocrinology. 2013 Jun;154(6):2200-7. doi: 10.1210/en.2012-2230. Epub 2013 Apr 1. Endocrinology. 2013. PMID: 23546600 Free PMC article.
-
Gonadotropin-regulated testicular RNA helicase (GRTH/DDX25) gene: cell-specific expression and transcriptional regulation by androgen in transgenic mouse testis.J Cell Biochem. 2010 Apr 15;109(6):1142-7. doi: 10.1002/jcb.22493. J Cell Biochem. 2010. PMID: 20108248
-
Gonadotropin-regulated testicular RNA helicase (GRTH/DDX25): a multifunctional protein essential for spermatogenesis.J Androl. 2010 Jan-Feb;31(1):45-52. doi: 10.2164/jandrol.109.008219. Epub 2009 Oct 29. J Androl. 2010. PMID: 19875492 Free PMC article. Review.
-
Gonadotropin-regulated testicular helicase (GRTH/DDX25): an essential regulator of spermatogenesis.Trends Endocrinol Metab. 2007 Oct;18(8):314-20. doi: 10.1016/j.tem.2007.09.001. Epub 2007 Sep 21. Trends Endocrinol Metab. 2007. PMID: 17889551 Review.
Cited by
-
Androgen-induced activation of gonadotropin-regulated testicular RNA helicase (GRTH/Ddx25) transcription: essential role of a nonclassical androgen response element half-site.Mol Cell Biol. 2012 Apr;32(8):1566-80. doi: 10.1128/MCB.06002-11. Epub 2012 Feb 13. Mol Cell Biol. 2012. PMID: 22331472 Free PMC article.
-
Effect of Early Calf-Hood Nutrition on the Transcriptional Regulation of the Hypothalamic-Pituitary-Testicular axis in Holstein-Friesian Bull Calves.Sci Rep. 2018 Nov 8;8(1):16577. doi: 10.1038/s41598-018-34611-4. Sci Rep. 2018. PMID: 30409985 Free PMC article.
-
Characterization of the Phosphorylation Site of GRTH/DDX25 and Protein Kinase A Binding Interface Provides Structural Basis for the Design of a Non-Hormonal Male Contraceptive.Sci Rep. 2019 Apr 30;9(1):6705. doi: 10.1038/s41598-019-42857-9. Sci Rep. 2019. PMID: 31040297 Free PMC article.
-
Germ Cell Nuclear Factor (GCNF/RTR) Regulates Transcription of Gonadotropin-Regulated Testicular RNA Helicase (GRTH/DDX25) in Testicular Germ Cells--The Androgen Connection.Mol Endocrinol. 2015 Dec;29(12):1792-804. doi: 10.1210/me.2015-1198. Epub 2015 Oct 20. Mol Endocrinol. 2015. PMID: 26484580 Free PMC article.
-
Role of gonadotropin regulated testicular RNA helicase (GRTH/Ddx25) on polysomal associated mRNAs in mouse testis.PLoS One. 2012;7(3):e32470. doi: 10.1371/journal.pone.0032470. Epub 2012 Mar 30. PLoS One. 2012. PMID: 22479328 Free PMC article.
References
-
- Tang P. Z., Tsai-Morris C. H., Dufau M. L. (1999) J. Biol. Chem. 274, 37932–37940 - PubMed
-
- Sheng Y., Tsai-Morris C. H., Dufau M. L. (2003) J. Biol. Chem. 278, 27796–27803 - PubMed
-
- Haelens A., Verrijdt G., Schoenmakers E., Alen P., Peeters B., Rombauts W., Claessens F. (1999) Mol. Cell. Endocrinol. 153, 91–102 - PubMed
-
- Tsai-Morris C. H., Lei S., Jiang Q., Sheng Y., Dufau M. L. (2004) Gene 331, 83–94 - PubMed
-
- Villar J., Tsai-Morris C. H., Dufau M. L. (2010) FASEB J. 24, 833.26. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
