

Membrane-anchored serine proteases in vertebrate cell and developmental biology
- PMID: 21721945
- PMCID: PMC3391589
- DOI: 10.1146/annurev-cellbio-092910-154247
Membrane-anchored serine proteases in vertebrate cell and developmental biology
Abstract
Analysis of vertebrate genome sequences at the turn of the millennium revealed that a vastly larger repertoire of enzymes execute proteolytic cleavage reactions within the pericellular and extracellular environments than was anticipated from biochemical and molecular analysis. Most unexpected was the unveiling of an entire new family of structurally unique multidomain serine proteases that are anchored directly to the plasma membrane. Unlike secreted serine proteases, which function primarily in tissue repair, immunity, and nutrient uptake, these membrane-anchored serine proteases regulate fundamental cellular and developmental processes, including tissue morphogenesis, epithelial barrier function, ion and water transport, cellular iron export, and fertilization. Here the cellular and developmental biology of this fascinating new group of proteases is reviewed. Particularly highlighted is how the study of membrane-anchored serine proteases has expanded our knowledge of the range of physiological processes that require regulated proteolysis at the cell surface.
Figures
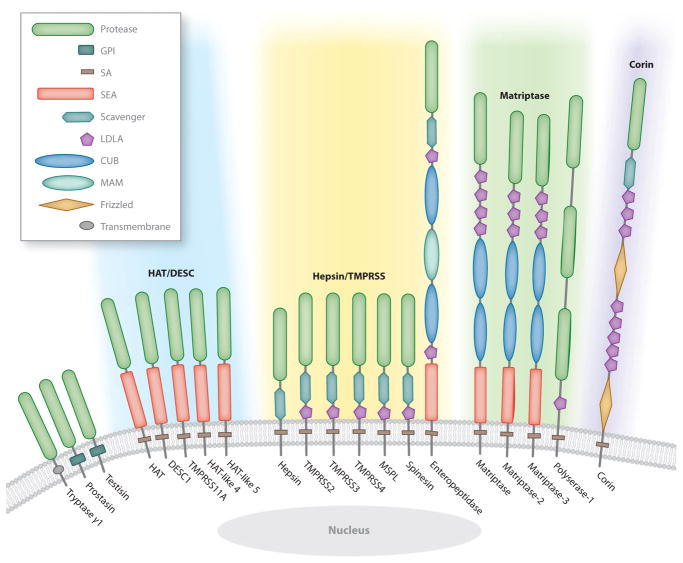

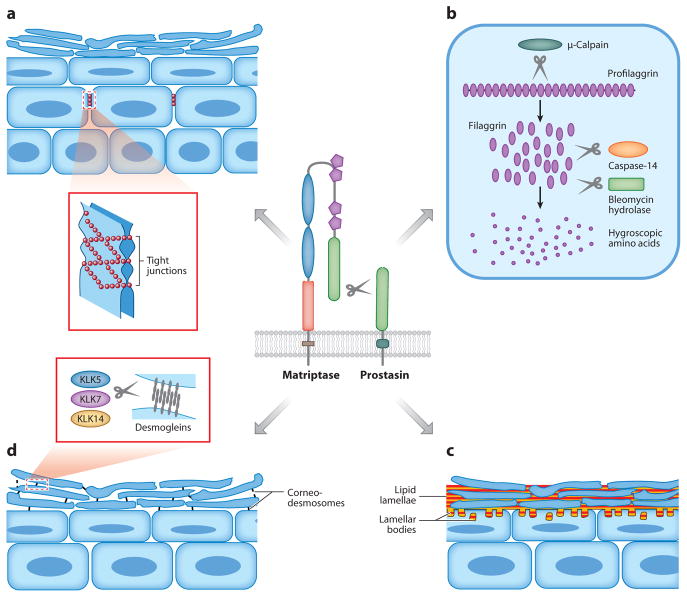
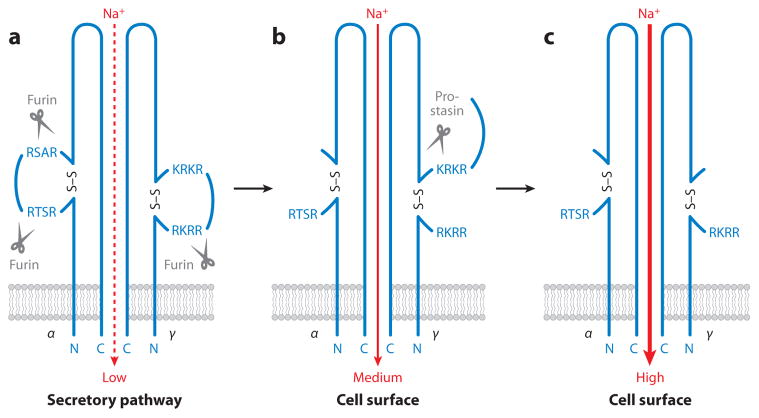
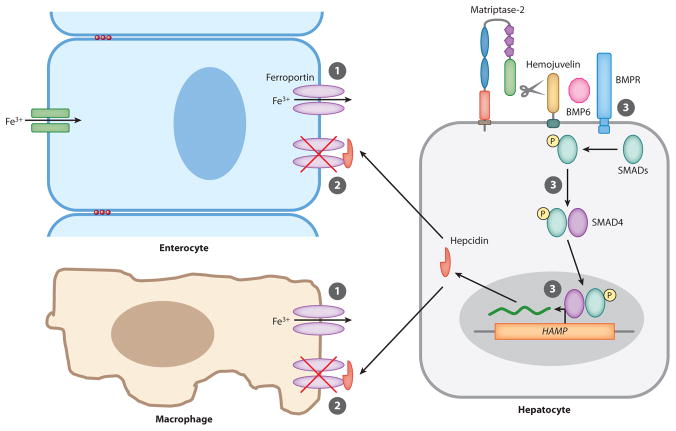
References
-
- Adham IM, Nayernia K, Engel W. Spermatozoa lacking acrosin protein show delayed fertilization. Mol Reprod Dev. 1997;46:370–76. - PubMed
-
- Alef T, Torres S, Hausser I, Metze D, Tursen U, et al. Ichthyosis, follicular atrophoderma, and hypotrichosis caused by mutations in ST14 is associated with impaired profilaggrin processing. J Investig Dermatol. 2009;129:862–69. - PubMed
-
- Andreasen D, Vuagniaux G, Fowler-Jaeger N, Hummler E, Rossier BC. Activation of epithelial sodium channels by mouse channel activating proteases (mCAP) expressed in Xenopus oocytes requires catalytic activity of mCAP3 and mCAP2 but not mCAP1. J Am Soc Nephrol. 2006;17:968–76. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
