

Xanthophyll cycle--a mechanism protecting plants against oxidative stress
- PMID: 21722416
- PMCID: PMC6837671
- DOI: 10.1179/174329211X13020951739938
Xanthophyll cycle--a mechanism protecting plants against oxidative stress
Abstract
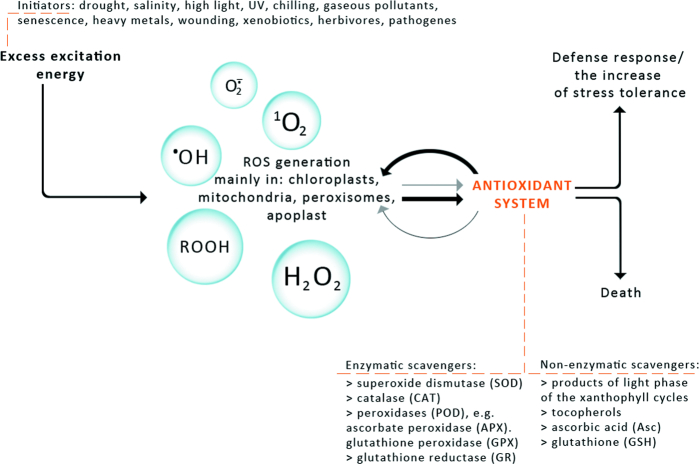
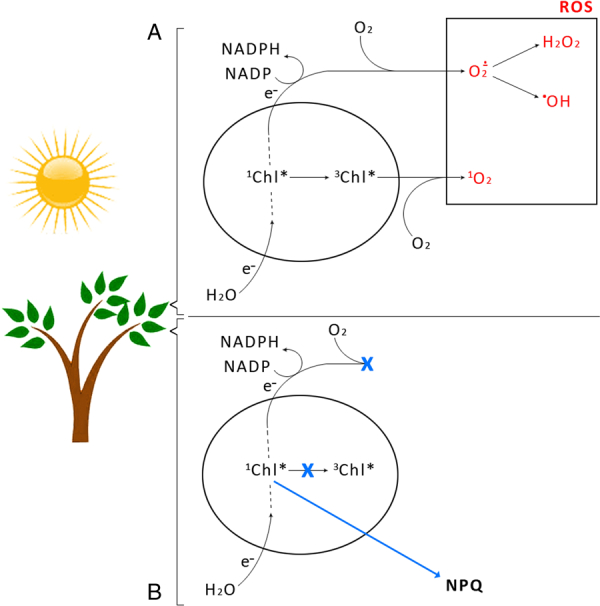
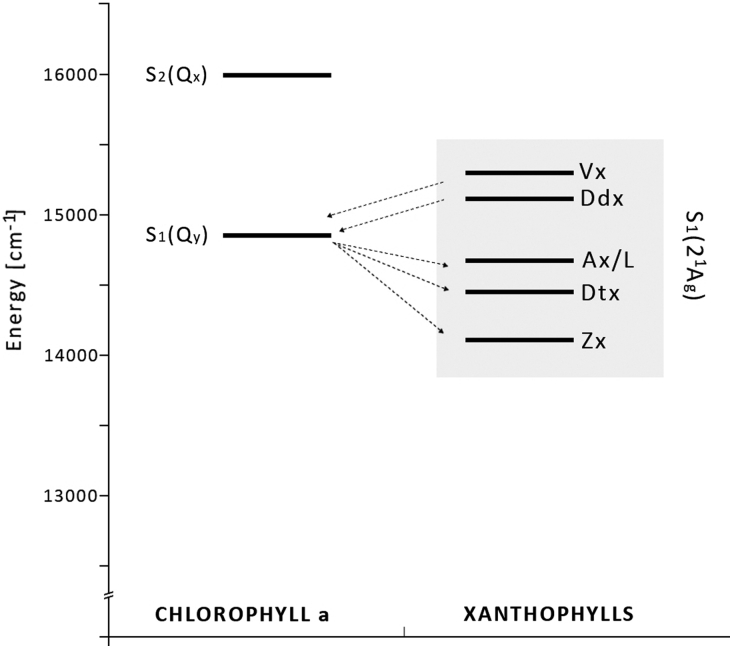
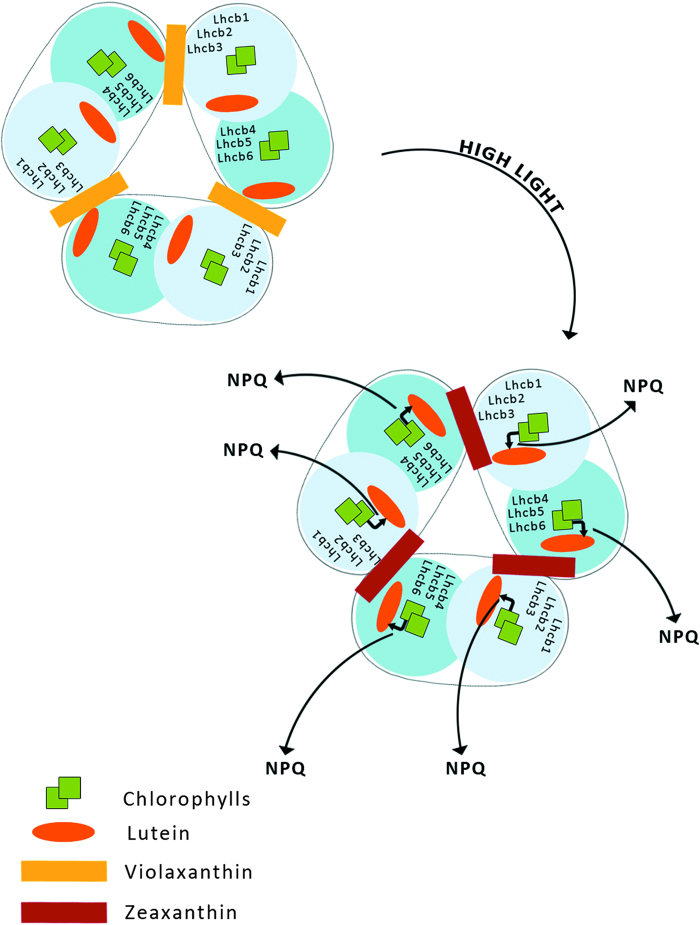
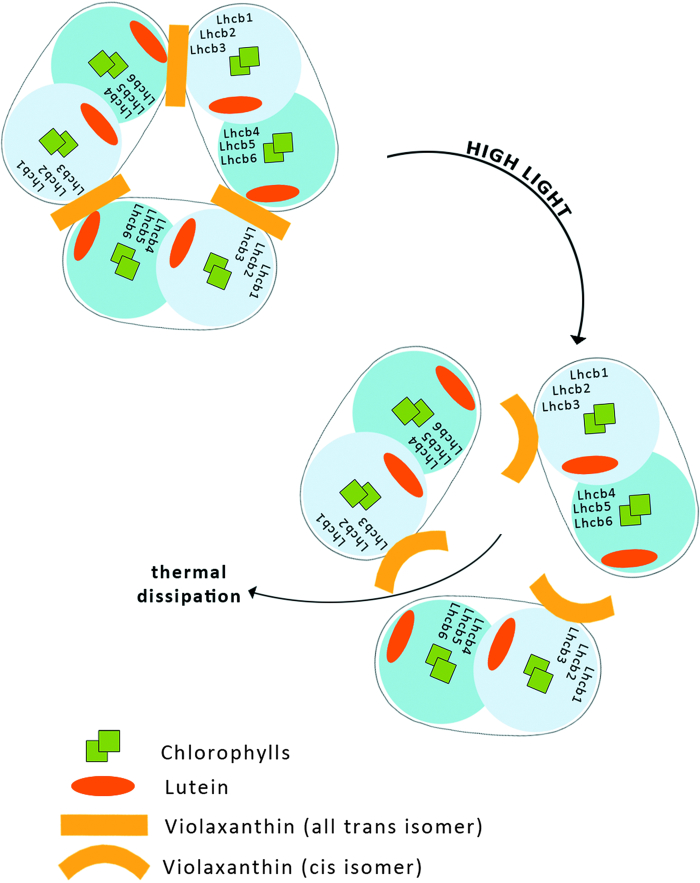
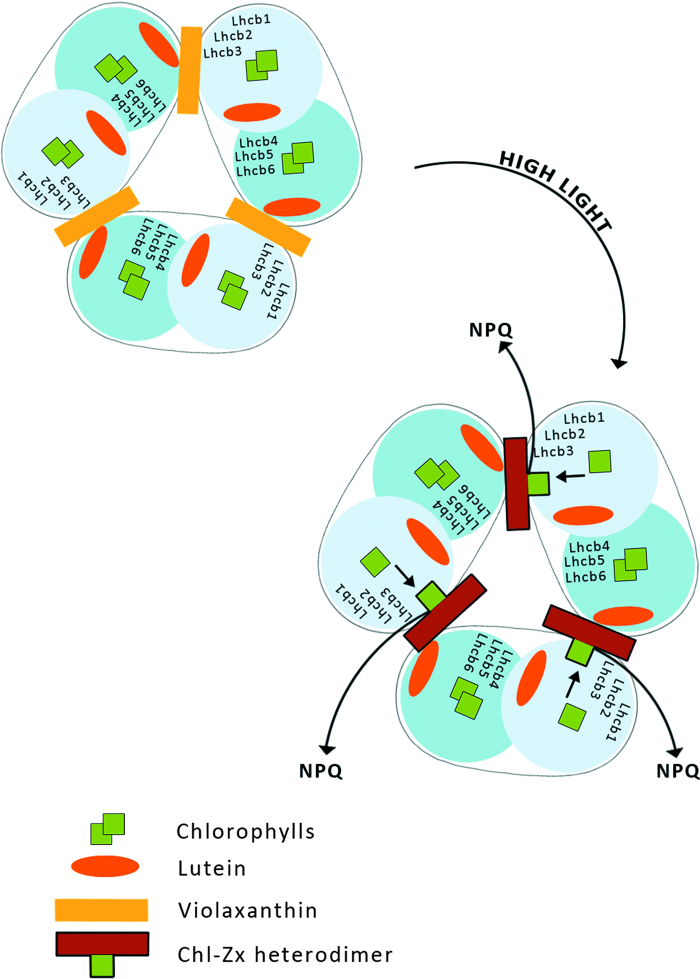
Six different xanthophyll cycles have been described in photosynthetic organisms. All of them protect the photosynthetic apparatus from photodamage caused by light-induced oxidative stress. Overexcitation conditions lead, in the chloroplast, to the over-reduction of the NADP pool and production of superoxide, which can subsequently be metabolized to hydrogen peroxide or a hydroxyl radical, other reactive oxygen species (ROS). On the other hand, overexcitation of photosystems leads to an increased lifetime of the chlorophyll excited state, increasing the probability of chlorophyll triplet formation which reacts with triplet oxygen forming single oxygen, another ROS. The products of the light-dependent phase of xanthophyll cycles play an important role in the protection against oxidative stress generated not only by an excess of light but also by other ROS-generating factors such as drought, chilling, heat, senescence, or salinity stress. Four, mainly hypothetical, mechanisms explaining the protective role of xanthophyll cycles in oxidative stress are presented. One of them is the direct quenching of overexcitation by products of the light phase of xanthophyll cycles and three others are based on the indirect participation of xanthophyll cycle carotenoids in the process of photoprotection. They include: (1) indirect quenching of overexcitation by aggregation-dependent light-harvesting complexes (LHCII) quenching; (2) light-driven mechanisms in LHCII; and (3) a model based on charge transfer quenching between Chl a and Zx. Moreover, results of the studies on the antioxidant properties of xanthophyll cycle pigments in model systems are also presented.
Figures
References
-
- Schafer FQ, Buettner GR. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic Biol Med 2001;30(11):1191–212. doi: 10.1016/S0891-5849(01)00480-4. PMID 11368918. - PubMed
-
- Lennon SV, Martin SJ, Cotter TG. Dose-dependent induction of apoptosis in human tumour cell lines by widely diverging stimuli. Cell Prolif 1991;24(2):203–14. doi: 10.1111/j.1365-2184.1991.tb01150.x. PMID 2009322. - PubMed
-
- Valko M, Morris H, Cronin MT. Metals, toxicity and oxidative stress. Curr Med Chem 2005;12(10):1161–208. doi: 10.2174/0929867053764635. PMID 15892631. - PubMed
-
- Puddu P, Puddu GM, Cravero E, Rosati M, Muscari A. The molecular sources of reactive oxygen species in hypertension. Blood Press 2008;17(2):70–7. - PubMed
-
- Rodriguez-Pallares J, Parga JA, Joglar B, Guerra MJ, Labandeira-Garcia JL. The mitochondrial ATP-sensitive potassium channel blocker 5-hydroxydecanoate inhibits toxicity of 6-hydroxydopamine on dopaminergic neurons. Neurotox Res 2009;15(1):82–95. Epub 2009 Feb 24. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources