

Structural basis for the cooperative DNA recognition by Smad4 MH1 dimers
- PMID: 21724602
- PMCID: PMC3185416
- DOI: 10.1093/nar/gkr500
Structural basis for the cooperative DNA recognition by Smad4 MH1 dimers
Abstract
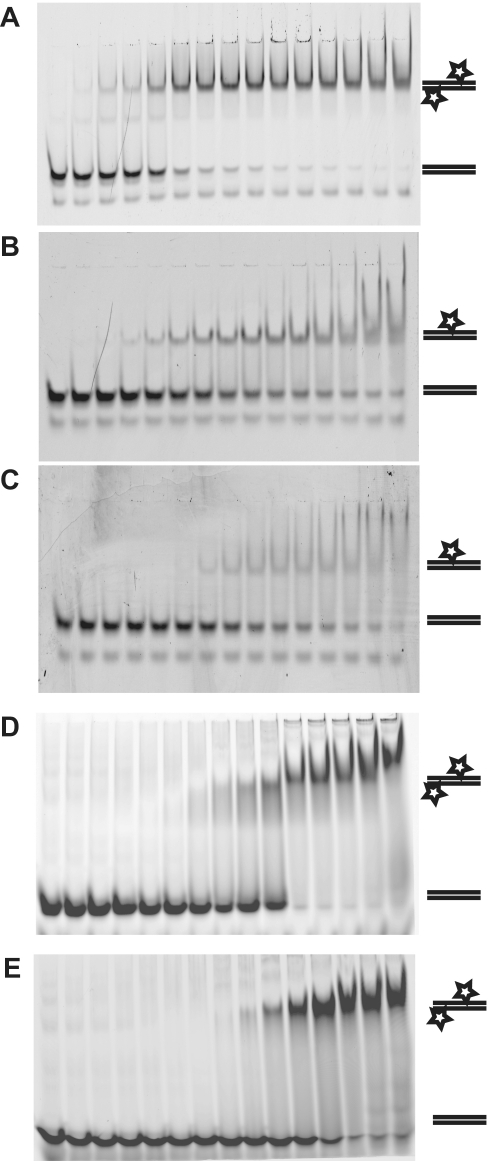
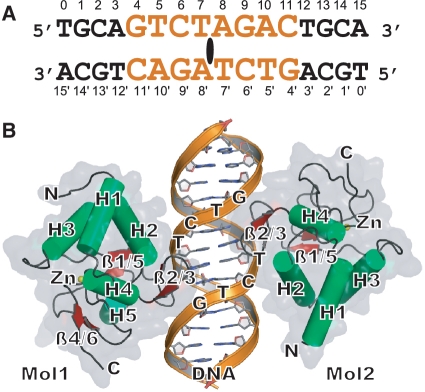
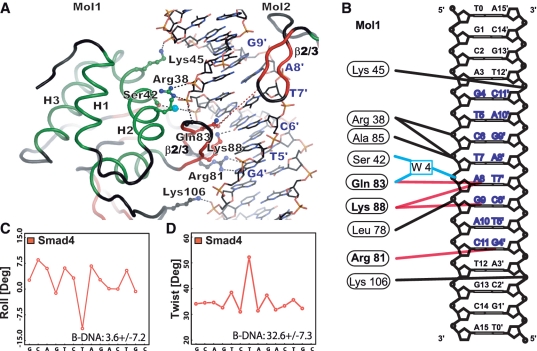
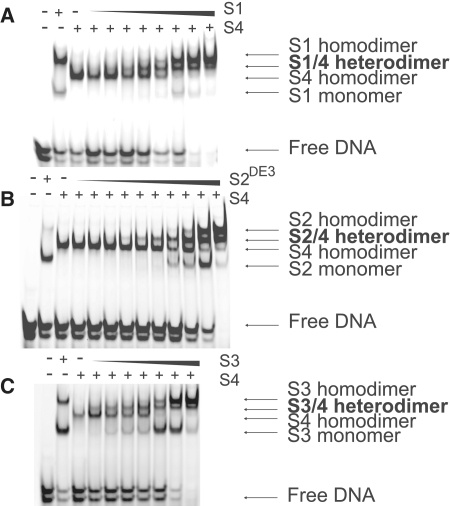
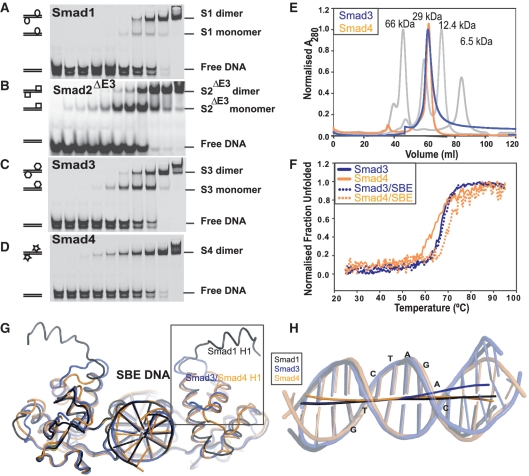
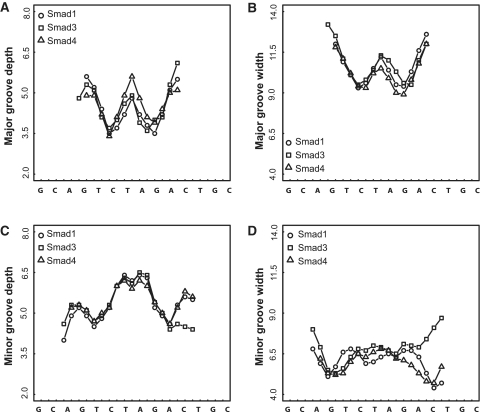
Smad proteins form multimeric complexes consisting of the 'common partner' Smad4 and receptor regulated R-Smads on clustered DNA binding sites. Deciphering how pathway specific Smad complexes multimerize on DNA to regulate gene expression is critical for a better understanding of the cis-regulatory logic of TGF-β and BMP signaling. To this end, we solved the crystal structure of the dimeric Smad4 MH1 domain bound to a palindromic Smad binding element. Surprisingly, the Smad4 MH1 forms a constitutive dimer on the SBE DNA without exhibiting any direct protein-protein interactions suggesting a DNA mediated indirect readout mechanism. However, the R-Smads Smad1, Smad2 and Smad3 homodimerize with substantially decreased efficiency despite pronounced structural similarities to Smad4. Therefore, intricate variations in the DNA structure induced by different Smads and/or variant energetic profiles likely contribute to their propensity to dimerize on DNA. Indeed, competitive binding assays revealed that the Smad4/R-Smad heterodimers predominate under equilibrium conditions while R-Smad homodimers are least favored. Together, we present the structural basis for DNA recognition by Smad4 and demonstrate that Smad4 constitutively homo- and heterodimerizes on DNA in contrast to its R-Smad partner proteins by a mechanism independent of direct protein contacts.
Figures
References
-
- Whitman M. Smads and early developmental signaling by the TGFbeta superfamily. Genes Dev. 1998;12:2445–2462. - PubMed
-
- Massagué J. TGF-beta signal transduction. Annu. Rev. Biochem. 1998;67:753–791. - PubMed
-
- Hoodless P, Haerry T, Abdollah S, Stapleton M, O’Connor M, Attisano L, Wrana J. MADR1, a MAD-related protein that functions in BMP2 signaling pathways. Cell. 1996;85:489–500. - PubMed
-
- Kawai S, Faucheu C, Gallea S, Spinella-Jaegle S, Atfi A, Baron R, Roman S. Mouse smad8 phosphorylation downstream of BMP receptors ALK-2, ALK-3, and ALK-6 induces its association with Smad4 and transcriptional activity. Biochem. Biophys. Res. Commun. 2000;271:682–687. - PubMed
-
- Macías-Silva M, Abdollah S, Hoodless P, Pirone R, Attisano L, Wrana J. MADR2 is a substrate of the TGFbeta receptor and its phosphorylation is required for nuclear accumulation and signaling. Cell. 1996;87:1215–1224. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
