

Prolonged exposure to GH impairs insulin signaling in the heart
- PMID: 21727153
- PMCID: PMC3746341
- DOI: 10.1530/JME-11-0066
Prolonged exposure to GH impairs insulin signaling in the heart
Abstract
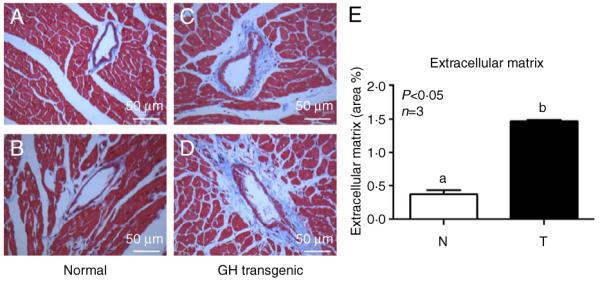
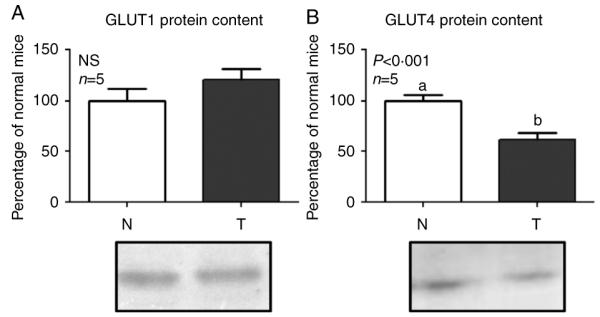
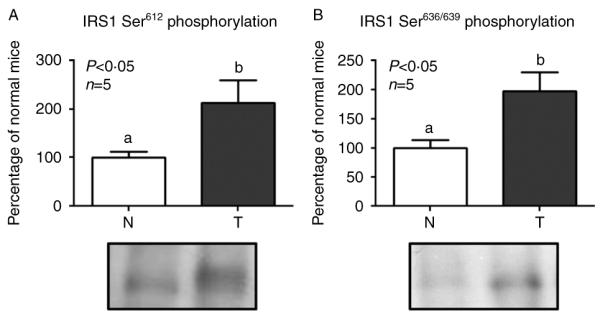
Acromegaly is associated with cardiac hypertrophy, which is believed to be a direct consequence of chronically elevated GH and IGF1. Given that insulin is important for cardiac growth and function, and considering that GH excess induces hyperinsulinemia, insulin resistance, and cardiac alterations, it is of interest to study insulin sensitivity in this tissue under chronic conditions of elevated GH. Transgenic mice overexpressing GH present cardiomegaly and perivascular and interstitial fibrosis in the heart. Mice received an insulin injection, the heart was removed after 2 min, and immunoblotting assays of tissue extracts were performed to evaluate the activation and abundance of insulin-signaling mediators. Insulin-induced tyrosine phosphorylation of the insulin receptor (IR) was conserved in transgenic mice, but the phosphorylation of IR substrate 1 (IRS1), its association with the regulatory subunit of the phosphatidylinositol 3-kinase (PI3K), and the phosphorylation of AKT were decreased. In addition, total content of the glucose transporter GLUT4 was reduced in transgenic mice. Insulin failed to induce the phosphorylation of the mammalian target of rapamycin (mTOR). However, transgenic mice displayed increased basal activation of the IR/IRS1/PI3K/AKT/mTOR and p38 signaling pathways along with higher serine phosphorylation of IRS1, which is recognized as an inhibitory modification. We conclude that GH-overexpressing mice exhibit basal activation of insulin signaling but decreased sensitivity to acute insulin stimulation at several signaling steps downstream of the IR in the heart. These alterations may be associated with the cardiac pathology observed in these animals.
Figures




References
-
- Abel ED, Kaulbach HC, Tian R, Hopkins JC, Duffy J, Doetschman T, Minnemann T, Boers ME, Hadro E, Oberste-Berghaus C, et al. Cardiac hypertrophy with preserved contractile function after selective deletion of GLUT4 from the heart. Journal of Clinical Investigation. 1999;104:1703–1714. (doi:10.1172/JCI7605) - PMC - PubMed
-
- Barbour LA, Mizanoor Rahman S, Gurevich I, Leitner JW, Fischer SJ, Roper MD, Knotts TA, Vo Y, McCurdy CE, Yakar S, et al. Increased P85alpha is a potent negative regulator of skeletal muscle insulin signaling and induces in vivo insulin resistance associated with growth hormone excess. Journal of Biological Chemistry. 2005;280:37489–37494. (doi:10.1074/jbc.M506967200) - PubMed
-
- Bartke A. Can growth hormone (GH) accelerate aging? Evidence from GH-transgenic mice Neuroendocrinology. 2003;78:210–216. (doi:10.1159/000073704) - PubMed
-
- Bartke A. Minireview: role of the growth hormone/insulin-like growth factor system in mammalian aging. Endocrinology. 2005;146:3718–3723. (doi:10.1210/en.2005-0411) - PubMed
-
- Belke DD, Betuing S, Tuttle MJ, Graveleau C, Young ME, Pham M, Zhang D, Cooksey RC, McClain DA, Litwin SE, et al. Insulin signaling coordinately regulates cardiac size, metabolism, and contractile protein isoform expression. Journal of Clinical Investigation. 2002;109:629–639. (doi:10.1172/JCI13946) - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
