

Fibrinogen-related proteins in ixodid ticks
- PMID: 21729260
- PMCID: PMC3141747
- DOI: 10.1186/1756-3305-4-127
Fibrinogen-related proteins in ixodid ticks
Abstract
Background: Fibrinogen-related proteins with lectin activity are believed to be part of the tick innate immune system. Several fibrinogen-related proteins have been described and characterised mainly on the basis of their cDNA sequences while direct biochemical evidence is missing. One of them, the haemolymph lectin Dorin M from the tick Ornithodoros moubata was isolated and characterised in more depth.
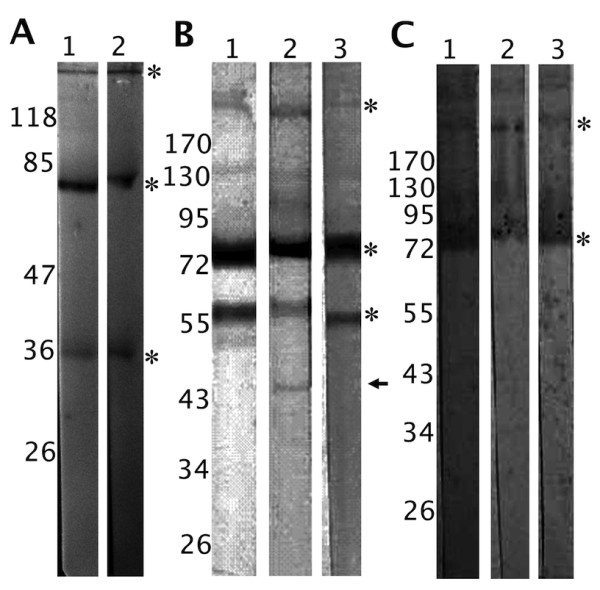
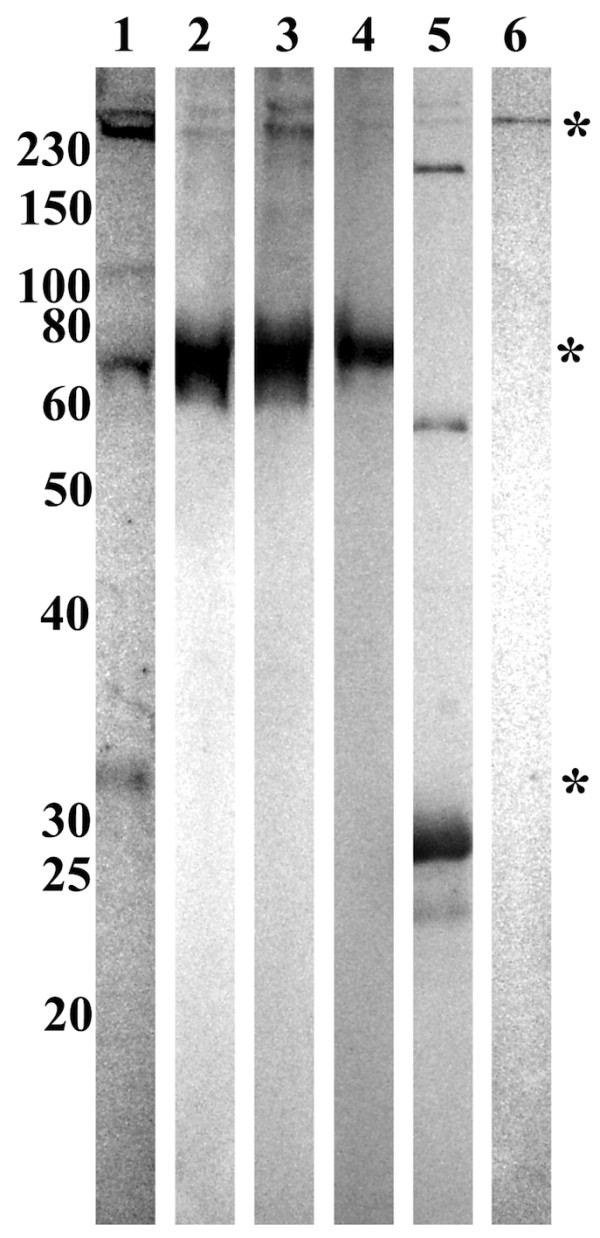
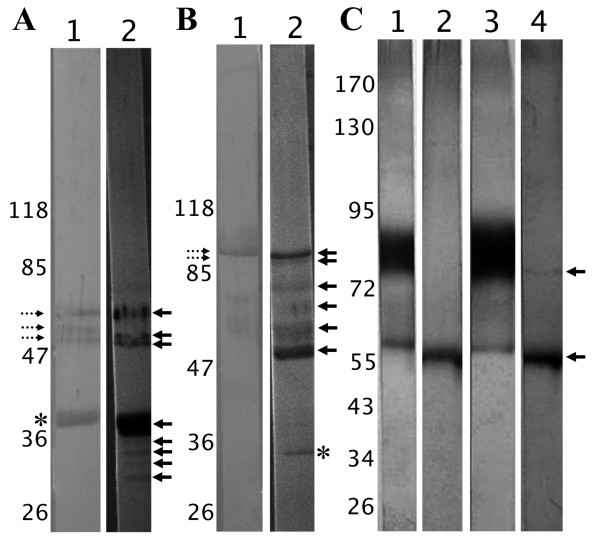
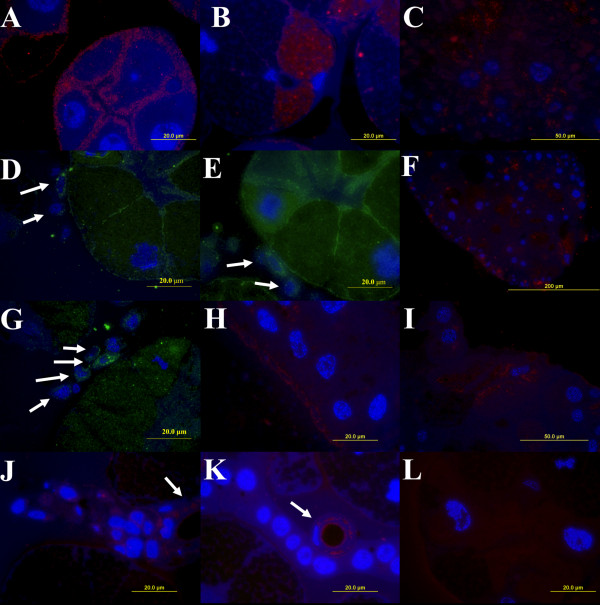
Results: Several fibrinogen-related proteins were detected in the haemolymph of ixodid ticks Dermacentor marginatus, Rhipicephalus appendiculatus, R. pulchellus, and R. sanguineus. These proteins were recognised by sera directed against the tick lectin Dorin M and the haemagglutination activity of the ticks R. appendiculatus and D. marginatus. Cross-reactivity of the identified proteins with antibodies against the fibrinogen domain of the human ficolin was also shown. The carbohydrate-binding ability of tick haemolymph was confirmed by haemagglutination activity assays, and this activity was shown to be inhibited by neuraminic acid and sialylated glycoproteins as well as by N-acetylated hexosamines. The fibrinogen-related proteins were shown to be glycosylated and they were localised in salivary glands, midguts, and haemocytes of D. marginatus. Hemelipoglycoprotein was also recognised by sera directed against the fibrinogen-related proteins in all three Rhipicephalus species as well as in D. marginatus. However, this protein does not contain the fibrinogen domain and thus, the binding possibly results from the structure similarity between hemelipoglycoprotein and the fibrinogen domain.
Conclusions: The presence of fibrinogen-related proteins was shown in the haemolymph of four tick species in high abundance. Reactivity of antibodies directed against ficolin or fibrinogen-related proteins with proteins which do not contain the fibrinogen domain points out the importance of sequence analysis of the identified proteins in further studies. Previously observed expression of fibrinogen-related proteins in haemocytes together with the results of this study suggest involvement of fibrinogen-related proteins in tick immunity processes. Thus, they have potential as targets for anti-tick vaccines and as antimicrobial proteins in pharmacology. Research on fibrinogen-related proteins could reveal further details of tick innate immunity processes.
Figures
References
-
- Sonenshine DE. Biology of ticks. Vol. 2. New York: Oxford University Press; 1993.
-
- Piesman J, Gage KL. In: The biology of disease vectors. Beaty BJ, Marquardt WC, editor. Niwot, CO: University of Colorado Press; 1996. Ticks and mites and the agents they transmit; pp. 160–174.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
