

Glioma stem cell proliferation and tumor growth are promoted by nitric oxide synthase-2
- PMID: 21729780
- PMCID: PMC3144745
- DOI: 10.1016/j.cell.2011.06.006
Glioma stem cell proliferation and tumor growth are promoted by nitric oxide synthase-2
Abstract
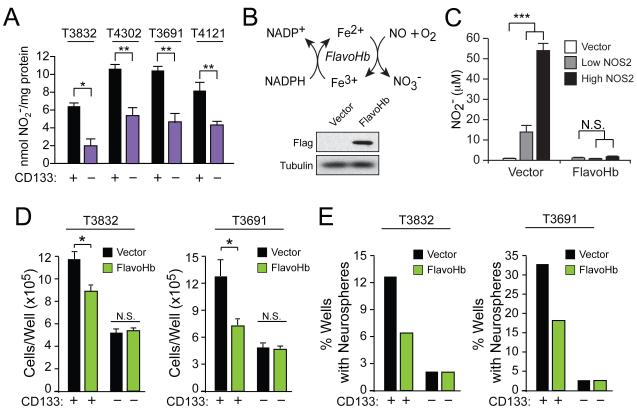
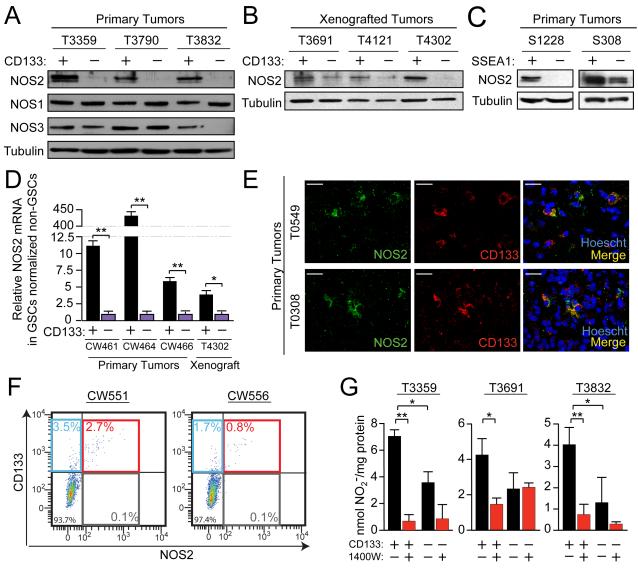
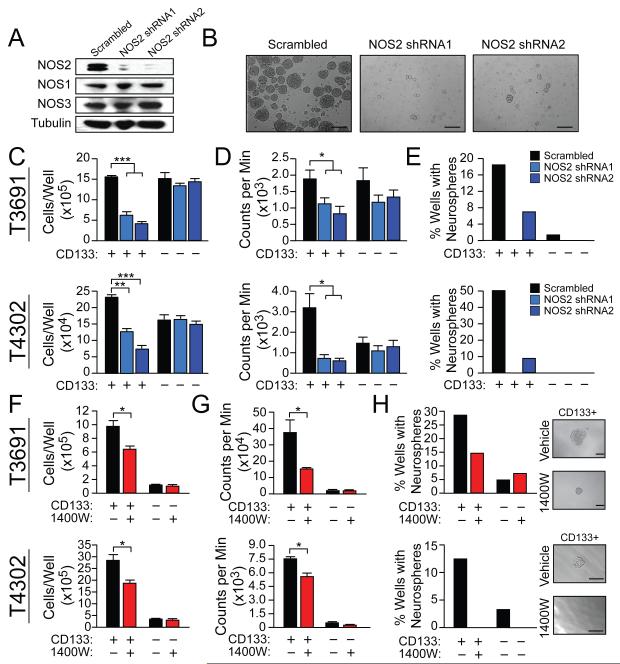
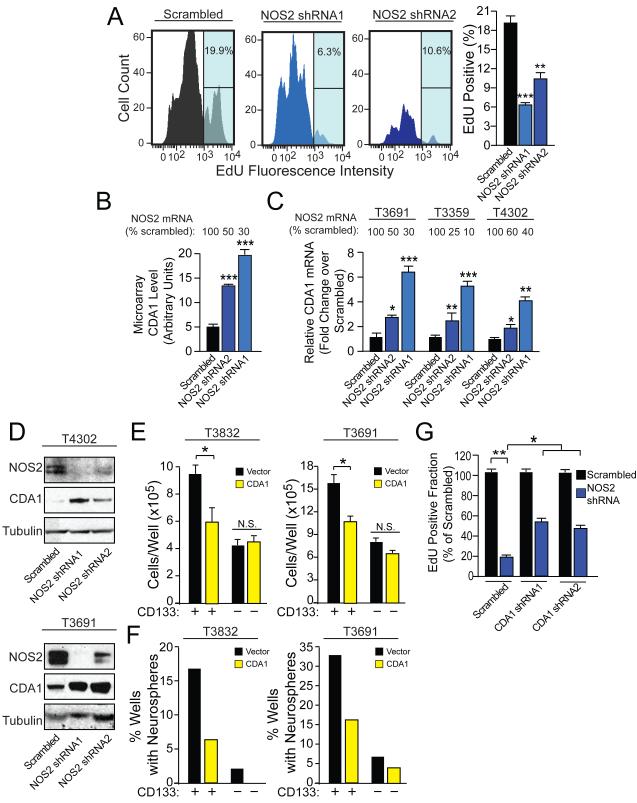
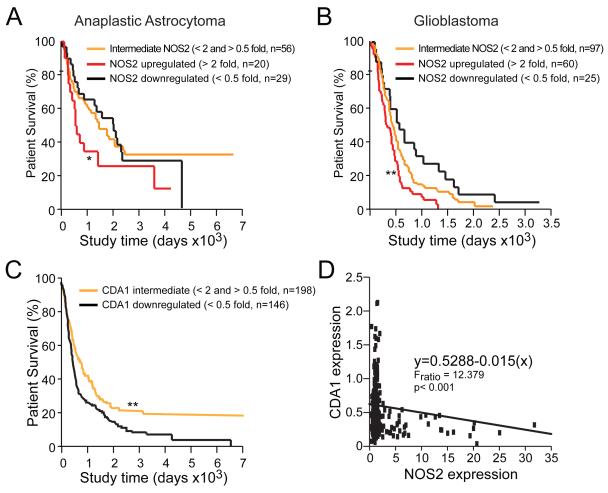
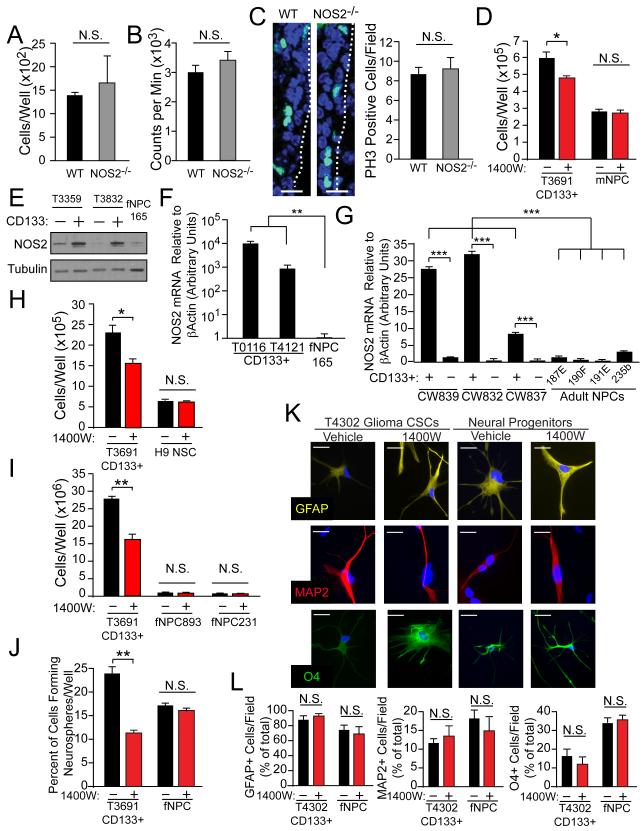
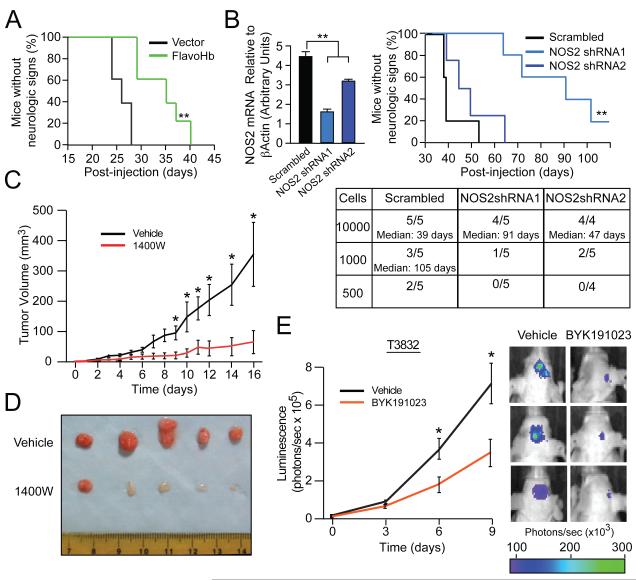
Malignant gliomas are aggressive brain tumors with limited therapeutic options, and improvements in treatment require a deeper molecular understanding of this disease. As in other cancers, recent studies have identified highly tumorigenic subpopulations within malignant gliomas, known generally as cancer stem cells. Here, we demonstrate that glioma stem cells (GSCs) produce nitric oxide via elevated nitric oxide synthase-2 (NOS2) expression. GSCs depend on NOS2 activity for growth and tumorigenicity, distinguishing them from non-GSCs and normal neural progenitors. Gene expression profiling identified many NOS2-regulated genes, including the cell-cycle inhibitor cell division autoantigen-1 (CDA1). Further, high NOS2 expression correlates with decreased survival in human glioma patients, and NOS2 inhibition slows glioma growth in a murine intracranial model. These data provide insight into how GSCs are mechanistically distinct from their less tumorigenic counterparts and suggest that NOS2 inhibition may be an efficacious approach to treating this devastating disease.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Alexander JH, Reynolds HR, Stebbins AL, Dzavik V, Harrington RA, Van de Werf F, Hochman JS. Effect of tilarginine acetate in patients with acute myocardial infarction and cardiogenic shock: the TRIUMPH randomized controlled trial. JAMA. 2007;297:1657–1666. - PubMed
-
- Androutsellis-Theotokis A, Leker RR, Soldner F, Hoeppner DJ, Ravin R, Poser SW, Rueger MA, Bae SK, Kittappa R, McKay RD. Notch signalling regulates stem cell numbers in vitro and in vivo. Nature. 2006;442:823–826. - PubMed
-
- Avontuur JA, Nolthenius R.P. Tutein, van Bodegom JW, Bruining HA. Prolonged inhibition of nitric oxide synthesis in severe septic shock: a clinical study. Crit Care Med. 1998;26:660–667. - PubMed
-
- Bakshi A, Nag TC, Wadhwa S, Mahapatra AK, Sarkar C. The expression of nitric oxide synthases in human brain tumours and peritumoral areas. J Neurol Sci. 1998;155:196–203. - PubMed
-
- Bao S, Wu Q, McLendon RE, Hao Y, Shi Q, Hjelmeland AB, Dewhirst MW, Bigner DD, Rich JN. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature. 2006a;444:756–760. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- P50 NS020023/NS/NINDS NIH HHS/United States
- R01 CA116659/CA/NCI NIH HHS/United States
- K08 CA101954/CA/NCI NIH HHS/United States
- R01 CA151522/CA/NCI NIH HHS/United States
- CA112958/CA/NCI NIH HHS/United States
- R01 HL059130/HL/NHLBI NIH HHS/United States
- HL091876/HL/NHLBI NIH HHS/United States
- R01 CA129958/CA/NCI NIH HHS/United States
- CA101954/CA/NCI NIH HHS/United States
- R01 CA116257/CA/NCI NIH HHS/United States
- CA137443/CA/NCI NIH HHS/United States
- CA142159/CA/NCI NIH HHS/United States
- R01 HL091876/HL/NHLBI NIH HHS/United States
- CA128269/CA/NCI NIH HHS/United States
- P50 CA108786/CA/NCI NIH HHS/United States
- CA116257/CA/NCI NIH HHS/United States
- NS20023/NS/NINDS NIH HHS/United States
- HL059130/HL/NHLBI NIH HHS/United States
- R01 HL095463/HL/NHLBI NIH HHS/United States
- R01 NS075345/NS/NINDS NIH HHS/United States
- F30NS063496/NS/NINDS NIH HHS/United States
- R42 CA128269/CA/NCI NIH HHS/United States
- CA108786/CA/NCI NIH HHS/United States
- R01 NS052534/NS/NINDS NIH HHS/United States
- F30 NS063496/NS/NINDS NIH HHS/United States
- CA154130/CA/NCI NIH HHS/United States
- CA116659/CA/NCI NIH HHS/United States
- HLO95463/PHS HHS/United States
- U01 CA137443/CA/NCI NIH HHS/United States
- UL1 TR000439/TR/NCATS NIH HHS/United States
- NS063971/NS/NINDS NIH HHS/United States
- CA151522/CA/NCI NIH HHS/United States
- F32 CA142159/CA/NCI NIH HHS/United States
- R01 NS063971/NS/NINDS NIH HHS/United States
- R01 NS054276/NS/NINDS NIH HHS/United States
- T32 GM007250/GM/NIGMS NIH HHS/United States
- R01 CA154130/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
