

Ezrin-mediated apical integrity is required for intestinal homeostasis
- PMID: 21730140
- PMCID: PMC3141968
- DOI: 10.1073/pnas.1103418108
Ezrin-mediated apical integrity is required for intestinal homeostasis
Abstract
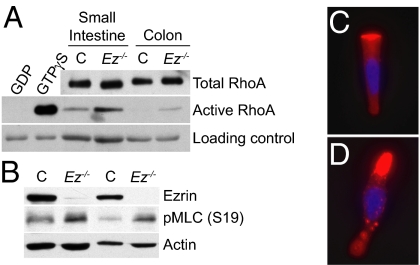
Individual cell types are defined by architecturally and functionally specialized cortical domains. The Ezrin, Radixin, and Moesin (ERM) proteins play a major role in organizing cortical domains by assembling membrane protein complexes and linking them to the cortical actin cytoskeleton. Many studies have focused on the individual roles of the ERM proteins in stabilizing the membrane-cytoskeleton interface, controlling the distribution and function of apical membrane complexes, regulating the small GTPase Rho, or establishing cell-cell junctions. We previously found that deletion of the mouse Ezrin gene yields severe defects in apical integrity throughout the developing intestinal epithelium, resulting in incomplete villus morphogenesis and neonatal death. However, the molecular function of Ezrin in building the apical surface of the intestinal epithelium was not clear. By deleting Ezrin in the adult mouse intestinal epithelium, we provide evidence that Ezrin performs multiple molecular functions that collaborate to build the functional apical surface of the intestinal epithelium in vivo. The loss of Ezrin-mediated apical integrity in the adult intestine yields severe morphological consequences during intestinal homeostasis, including defects in cell geometry, extrusion, junctional remodeling, and spindle orientation. Surprisingly, deletion of Ezrin either before or after villus morphogenesis yields villus fusion, revealing a previously unrecognized step in intestinal homeostasis. Our studies indicate that the function of Ezrin in building and maintaining the apical domain is essential not only for intestinal morphogenesis but also for homeostasis in the mature intestine.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Pearson MA, Reczek D, Bretscher A, Karplus PA. Structure of the ERM protein moesin reveals the FERM domain fold masked by an extended actin binding tail domain. Cell. 2000;101:259–270. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
