

Transcriptional activation of TINF2, a gene encoding the telomere-associated protein TIN2, by Sp1 and NF-κB factors
- PMID: 21731707
- PMCID: PMC3121743
- DOI: 10.1371/journal.pone.0021333
Transcriptional activation of TINF2, a gene encoding the telomere-associated protein TIN2, by Sp1 and NF-κB factors
Abstract
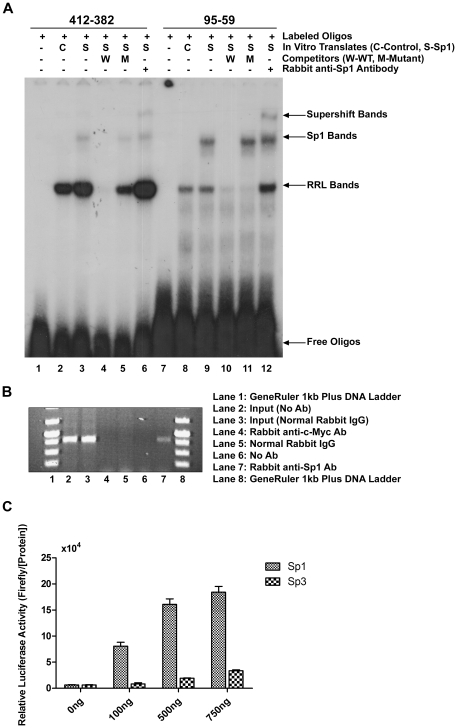
The expression of the telomere-associated protein TIN2 has been shown to be essential for early embryonic development in mice and for development of a variety of human malignancies. Recently, germ-line mutations in TINF2, which encodes for the TIN2 protein, have been identified in a number of patients with bone-marrow failure syndromes. Yet, the molecular mechanisms that regulate TINF2 expression are largely unknown. To elucidate the mechanisms involved in human TINF2 regulation, we cloned a 2.7 kb genomic DNA fragment containing the putative promoter region and, through deletion analysis, identified a 406 bp region that functions as a minimal promoter. This promoter proximal region is predicted to contain several putative Sp1 and NF-κB binding sites based on bioinformatic analysis. Direct binding of the Sp1 and NF-κB transcription factors to the TIN2 promoter sequence was demonstrated by electrophoretic mobility shift assay (EMSA) and/or chromatin immunoprecipitation (ChIP) assays. Transfection of a plasmid carrying the Sp1 transcription factor into Sp-deficient SL2 cells strongly activated TIN2 promoter-driven luciferase reporter expression. Similarly, the NF-κB molecules p50 and p65 were found to strongly activate luciferase expression in NF-κB knockout MEFs. Mutating the predicted transcription factor binding sites effectively reduced TIN2 promoter activity. Various known chemical inhibitors of Sp1 and NF-κB could also strongly inhibit TIN2 transcriptional activity. Collectively, our results demonstrate the important roles that Sp1 and NF-κB play in regulating the expression of the human telomere-binding protein TIN2, which can shed important light on its possible role in causing various forms of human diseases and cancers.
Conflict of interest statement
Figures




Similar articles
-
Characterization of the human intestinal CD98 promoter and its regulation by interferon-gamma.Am J Physiol Gastrointest Liver Physiol. 2007 Feb;292(2):G535-45. doi: 10.1152/ajpgi.00385.2006. Epub 2006 Oct 5. Am J Physiol Gastrointest Liver Physiol. 2007. PMID: 17023546
-
NFkappaB and Sp1 elements are necessary for maximal transcription of toll-like receptor 2 induced by Mycobacterium avium.J Immunol. 2001 Dec 15;167(12):6924-32. doi: 10.4049/jimmunol.167.12.6924. J Immunol. 2001. PMID: 11739511
-
Transcriptional regulation mechanisms of hypoxia-induced neuroglobin gene expression.Biochem J. 2012 Apr 1;443(1):153-64. doi: 10.1042/BJ20111856. Biochem J. 2012. PMID: 22239089
-
NF-κB, Sp1 and NF-Y as transcriptional regulators of human SND1 gene.Biochimie. 2013 Apr;95(4):735-42. doi: 10.1016/j.biochi.2012.10.029. Epub 2012 Nov 15. Biochimie. 2013. PMID: 23160072
-
Activation-dependent transcriptional regulation of the human Fas promoter requires NF-kappaB p50-p65 recruitment.Mol Cell Biol. 1999 Mar;19(3):2098-108. doi: 10.1128/MCB.19.3.2098. Mol Cell Biol. 1999. PMID: 10022897 Free PMC article.
Cited by
-
Identifying potential novel insights for COVID-19 pathogenesis and therapeutics using an integrated bioinformatics analysis of host transcriptome.Int J Biol Macromol. 2022 Jan 1;194:770-780. doi: 10.1016/j.ijbiomac.2021.11.124. Epub 2021 Nov 24. Int J Biol Macromol. 2022. PMID: 34826456 Free PMC article.
-
Transcriptional activation of Mina by Sp1/3 factors.PLoS One. 2013 Dec 4;8(12):e80638. doi: 10.1371/journal.pone.0080638. eCollection 2013. PLoS One. 2013. PMID: 24324617 Free PMC article.
-
Study of differential effects of TGF-beta3/BMP2 on chondrogenesis in MSC cells by gene microarray data analysis.Mol Cell Biochem. 2014 Jan;385(1-2):191-8. doi: 10.1007/s11010-013-1827-z. Epub 2013 Nov 7. Mol Cell Biochem. 2014. PMID: 24198161
References
-
- de Lange T. Shelterin: the protein complex that shapes and safeguards human telomeres. Genes Dev. 2005;19:2100–2110. - PubMed
-
- Chong L, van Steensel B, Broccoli D, Erdjument-Bromage H, Hanish J, et al. A human telomeric protein. Science. 1995;270:1663–1667. - PubMed
-
- Bilaud T, Brun C, Ancelin K, Koering CE, Laroche T, et al. Telomeric localization of TRF2, a novel human telobox protein. Nat Genet. 1997;17:236–239. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
