

Evolution of ruminant headgear: a review
- PMID: 21733893
- PMCID: PMC3151718
- DOI: 10.1098/rspb.2011.0938
Evolution of ruminant headgear: a review
Abstract
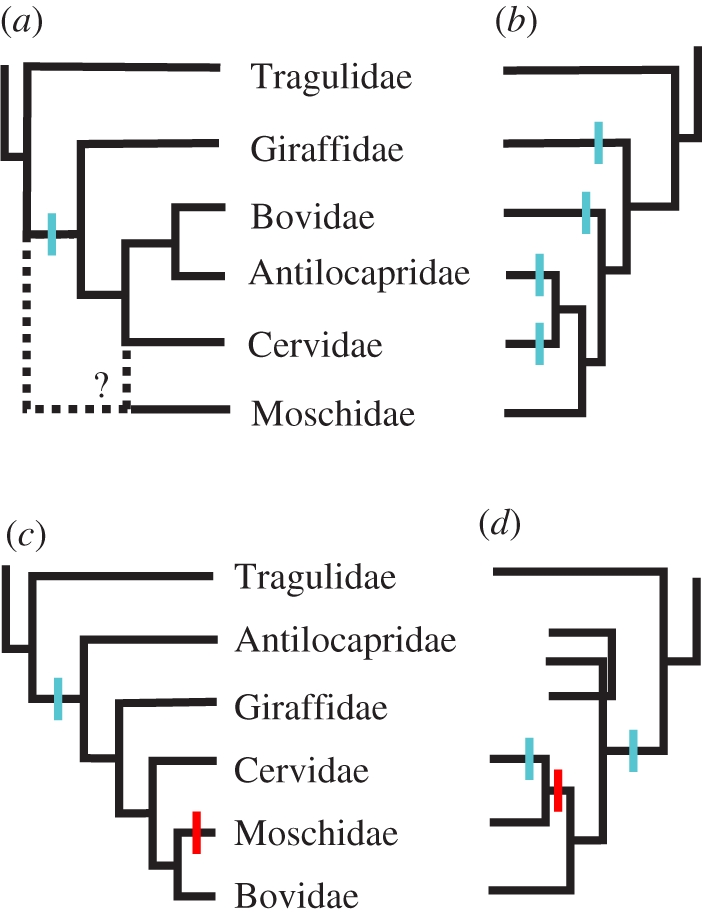
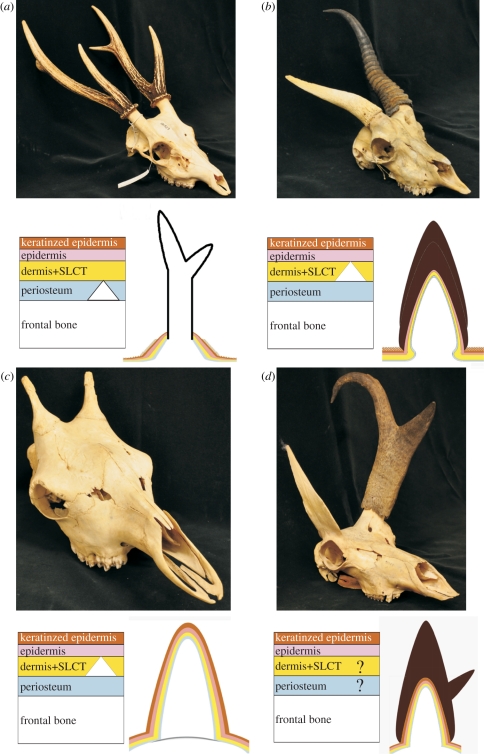
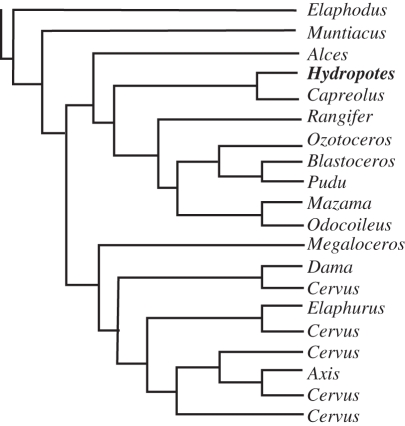
The horns, ossicones and antlers of ruminants are familiar and diverse examples of cranial appendages. We collectively term ruminant cranial appendages 'headgear'; this includes four extant forms: antlers (in cervids), horns (in bovids), pronghorns (in pronghorn antelope) and ossicones (in giraffids). Headgear evolution remains an open and intriguing question because phylogenies (molecular and morphological), adult headgear structure and headgear development (where data are available) all suggest different pictures of ruminant evolution. We discuss what is known about the evolution of headgear, including the evidence motivating previous hypotheses of single versus multiple origins, and the implications of recent phylogenetic revisions for these hypotheses. Inclusion of developmental data is critical for progress on the question of headgear evolution, and we synthesize the scattered literature on this front. The areas most in need of attention are early development in general; pronghorn and ossicone development in particular; and histological study of fossil forms of headgear. An integrative study of headgear development and evolution may have ramifications beyond the fields of systematics and evolution. Researchers in organismal biology, as well as those in biomedical fields investigating skin, bone and regenerative medicine, may all benefit from insights produced by this line of research.
Figures
References
-
- Jarman P. J. 1974. The social organisation of antelope in relation to their ecology. Behaviour 48, 215–26710.1163/156853974X00345 (doi:10.1163/156853974X00345) - DOI - DOI
-
- Reeves R. R., Tracey S. 1980. Monodon monoceros Mamm. Spec. 127, 1–710.2307/3503952 (doi:10.2307/3503952) - DOI - DOI
-
- Schneider D. 1964. Insect antennae. Annu. Rev. Entomol. 9, 103–12210.1146/annurev.en.09.010164.000535 (doi:10.1146/annurev.en.09.010164.000535) - DOI - DOI
-
- Krieger J., Breer H. 1999. Olfactory reception in invertebrates. Science 286, 720–72310.1126/science.286.5440.720 (doi:10.1126/science.286.5440.720) - DOI - DOI - PubMed
-
- Fay F. H. 1985. Odobenus rosmarus. Mamm. Spec. 238, 1–710.2307/3503810 (doi:10.2307/3503810) - DOI - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources