

Mechanisms and time course of vocal learning and consolidation in the adult songbird
- PMID: 21734110
- PMCID: PMC3191835
- DOI: 10.1152/jn.00311.2011
Mechanisms and time course of vocal learning and consolidation in the adult songbird
Abstract
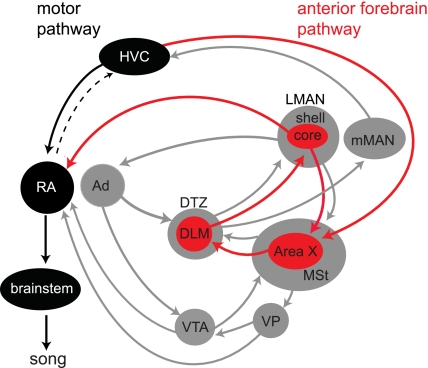
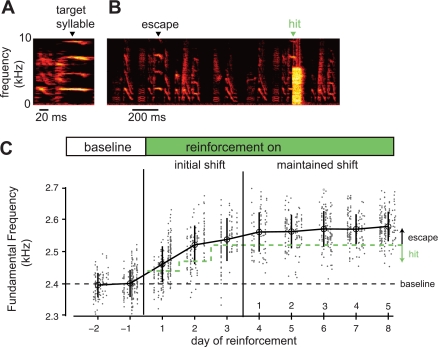
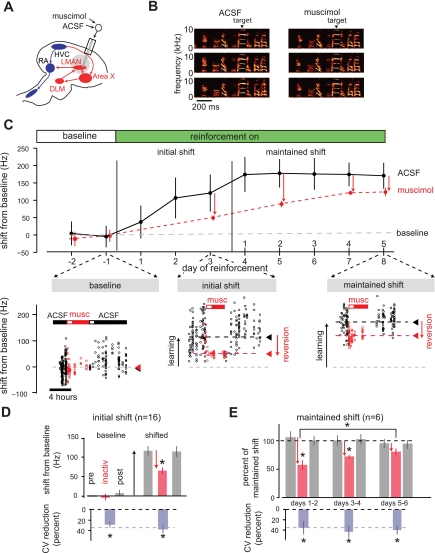
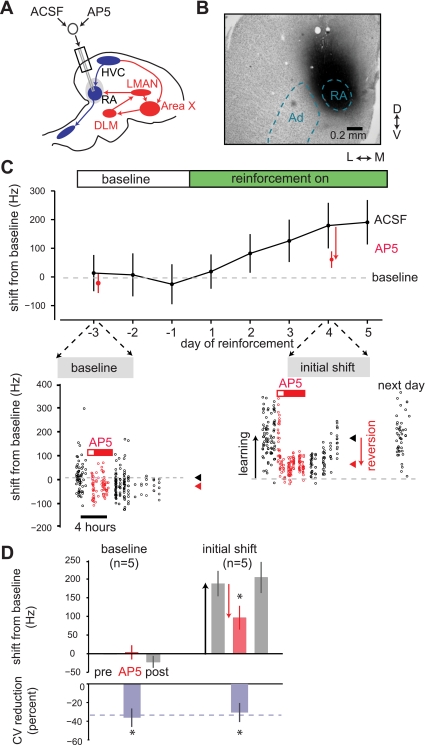
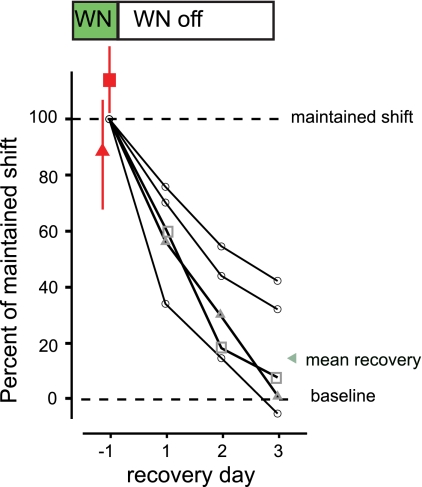
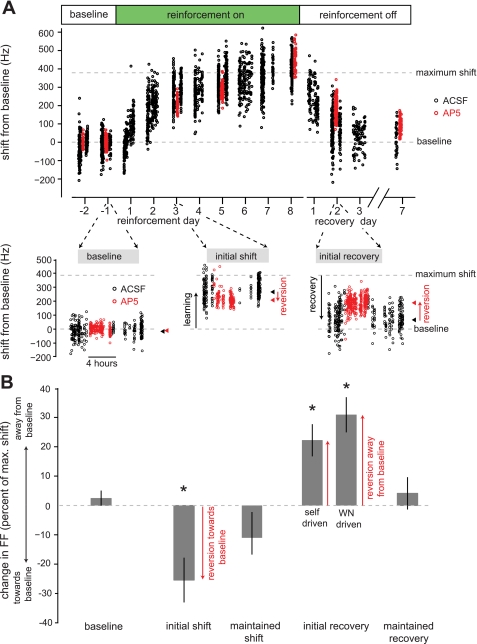
In songbirds, the basal ganglia outflow nucleus LMAN is a cortical analog that is required for several forms of song plasticity and learning. Moreover, in adults, inactivating LMAN can reverse the initial expression of learning driven via aversive reinforcement. In the present study, we investigated how LMAN contributes to both reinforcement-driven learning and a self-driven recovery process in adult Bengalese finches. We first drove changes in the fundamental frequency of targeted song syllables and compared the effects of inactivating LMAN with the effects of interfering with N-methyl-d-aspartate (NMDA) receptor-dependent transmission from LMAN to one of its principal targets, the song premotor nucleus RA. Inactivating LMAN and blocking NMDA receptors in RA caused indistinguishable reversions in the expression of learning, indicating that LMAN contributes to learning through NMDA receptor-mediated glutamatergic transmission to RA. We next assessed how LMAN's role evolves over time by maintaining learned changes to song while periodically inactivating LMAN. The expression of learning consolidated to become LMAN independent over multiple days, indicating that this form of consolidation is not completed over one night, as previously suggested, and instead may occur gradually during singing. Subsequent cessation of reinforcement was followed by a gradual self-driven recovery of original song structure, indicating that consolidation does not correspond with the lasting retention of changes to song. Finally, for self-driven recovery, as for reinforcement-driven learning, LMAN was required for the expression of initial, but not later, changes to song. Our results indicate that NMDA receptor-dependent transmission from LMAN to RA plays an essential role in the initial expression of two distinct forms of vocal learning and that this role gradually wanes over a multiday process of consolidation. The results support an emerging view that cortical-basal ganglia circuits can direct the initial expression of learning via top-down influences on primary motor circuitry.
Figures
Similar articles
-
Silent synapses in a thalamo-cortical circuit necessary for song learning in zebra finches.J Neurophysiol. 2005 Dec;94(6):3698-707. doi: 10.1152/jn.00282.2005. Epub 2005 Aug 17. J Neurophysiol. 2005. PMID: 16107531
-
Connections of a motor cortical region in zebra finches: relation to pathways for vocal learning.J Comp Neurol. 2000 May 1;420(2):244-60. J Comp Neurol. 2000. PMID: 10753310
-
Intrinsic and thalamic excitatory inputs onto songbird LMAN neurons differ in their pharmacological and temporal properties.J Neurophysiol. 1998 May;79(5):2615-28. doi: 10.1152/jn.1998.79.5.2615. J Neurophysiol. 1998. PMID: 9582233
-
Variability in action: Contributions of a songbird cortical-basal ganglia circuit to vocal motor learning and control.Neuroscience. 2015 Jun 18;296:39-47. doi: 10.1016/j.neuroscience.2014.10.010. Epub 2014 Oct 18. Neuroscience. 2015. PMID: 25445191 Review.
-
A hypothesis for basal ganglia-dependent reinforcement learning in the songbird.Neuroscience. 2011 Dec 15;198:152-70. doi: 10.1016/j.neuroscience.2011.09.069. Epub 2011 Oct 13. Neuroscience. 2011. PMID: 22015923 Free PMC article. Review.
Cited by
-
Discrete Evaluative and Premotor Circuits Enable Vocal Learning in Songbirds.Neuron. 2019 Nov 6;104(3):559-575.e6. doi: 10.1016/j.neuron.2019.07.025. Epub 2019 Aug 22. Neuron. 2019. PMID: 31447169 Free PMC article.
-
Interference and shaping in sensorimotor adaptations with rewards.PLoS Comput Biol. 2014 Jan;10(1):e1003377. doi: 10.1371/journal.pcbi.1003377. Epub 2014 Jan 9. PLoS Comput Biol. 2014. PMID: 24415925 Free PMC article.
-
Rules and mechanisms for efficient two-stage learning in neural circuits.Elife. 2017 Apr 4;6:e20944. doi: 10.7554/eLife.20944. Elife. 2017. PMID: 28374674 Free PMC article.
-
Generative models of birdsong learning link circadian fluctuations in song variability to changes in performance.PLoS Comput Biol. 2023 May 1;19(5):e1011051. doi: 10.1371/journal.pcbi.1011051. eCollection 2023 May. PLoS Comput Biol. 2023. PMID: 37126511 Free PMC article.
-
Variable but not random: temporal pattern coding in a songbird brain area necessary for song modification.J Neurophysiol. 2021 Feb 1;125(2):540-555. doi: 10.1152/jn.00034.2019. Epub 2020 Dec 9. J Neurophysiol. 2021. PMID: 33296616 Free PMC article.
References
-
- Amberg G, Lindefors N., 2nd Intracerebral microdialysis: mathematical studies of diffusion kinetics. J Pharmacol Methods 22: 157–183, 1989 - PubMed
-
- Appeltants D, Absil P, Balthazart J, Ball GF. Identification of the origin of catecholaminergic inputs to HVc in canaries by retrograde tract tracing combined with tyrosine hydroxylase immunocytochemistry. J Chem Neuroanat 18: 117–133, 2000 - PubMed
-
- Appeltants D, Ball GF, Balthazart J. The origin of catecholaminergic inputs to the song control nucleus RA in canaries. Neuroreport 13: 649–653, 2002 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources