

Inhibition of TET2-mediated conversion of 5-methylcytosine to 5-hydroxymethylcytosine disturbs erythroid and granulomonocytic differentiation of human hematopoietic progenitors
- PMID: 21734233
- PMCID: PMC3292425
- DOI: 10.1182/blood-2010-12-324707
Inhibition of TET2-mediated conversion of 5-methylcytosine to 5-hydroxymethylcytosine disturbs erythroid and granulomonocytic differentiation of human hematopoietic progenitors
Abstract
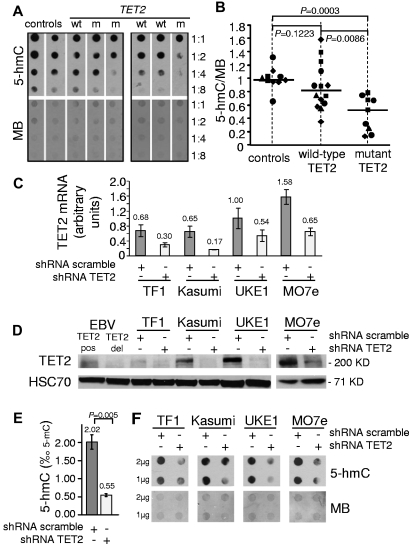
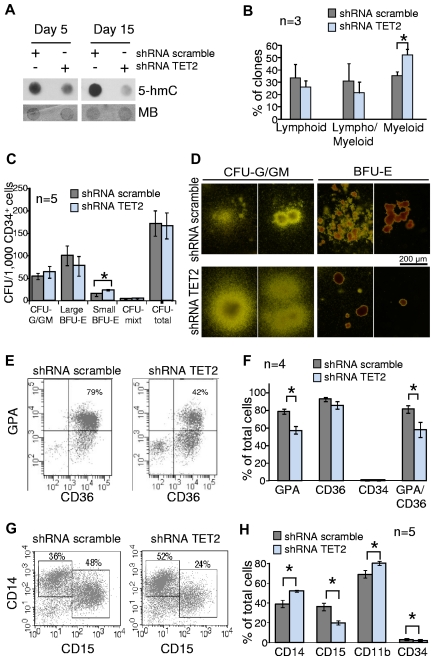
TET2 converts 5-methylcytosine to 5-hydroxymethylcytosine (5-hmC) in DNA and is frequently mutated in myeloid malignancies, including myeloproliferative neoplasms. Here we show that the level of 5-hmC is decreased in granulocyte DNA from myeloproliferative neoplasm patients with TET2 mutations compared with granulocyte DNA from healthy patients. Inhibition of TET2 by RNA interference decreases 5-hmC levels in both human leukemia cell lines and cord blood CD34(+) cells. These results confirm the enzymatic function of TET2 in human hematopoietic cells. Knockdown of TET2 in cord blood CD34(+) cells skews progenitor differentiation toward the granulomonocytic lineage at the expense of lymphoid and erythroid lineages. In addition, by monitoring in vitro granulomonocytic development we found a decreased granulocytic differentiation and an increase in monocytic cells. Our results indicate that TET2 disruption affects 5-hmC levels in human myeloid cells and participates in the pathogenesis of myeloid malignancies through the disturbance of myeloid differentiation.
Figures
Comment in
-
Leukaemia and lymphoma: The expansive reach of TET2.Nat Rev Cancer. 2011 Jul 22;11(8):535. doi: 10.1038/nrc3115. Nat Rev Cancer. 2011. PMID: 21779006 No abstract available.
References
-
- Ficz G, Branco MR, Seisenberger S, et al. Dynamic regulation of 5-hydroxymethylcytosine in mouse ES cells and during differentiation. Nature. 2011;473(7347):398–402. - PubMed
-
- Langemeijer SM, Kuiper RP, Berends M, et al. Acquired mutations in TET2 are common in myelodysplastic syndromes. Nat Genet. 2009;41(7):838–842. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
