

The striatum is highly susceptible to mitochondrial oxidative phosphorylation dysfunctions
- PMID: 21734281
- PMCID: PMC3175592
- DOI: 10.1523/JNEUROSCI.6223-10.2011
The striatum is highly susceptible to mitochondrial oxidative phosphorylation dysfunctions
Abstract
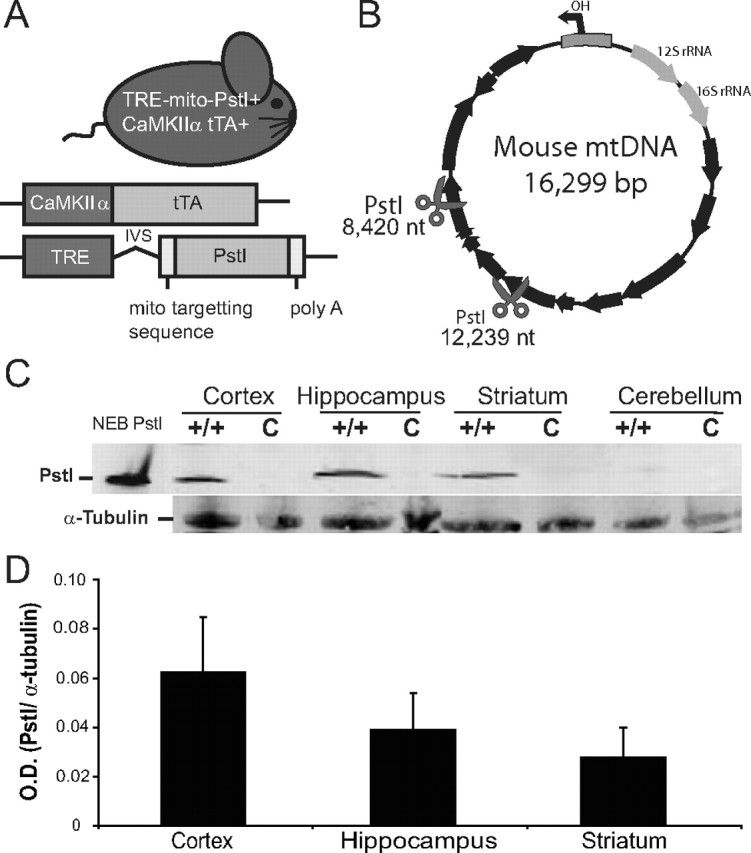
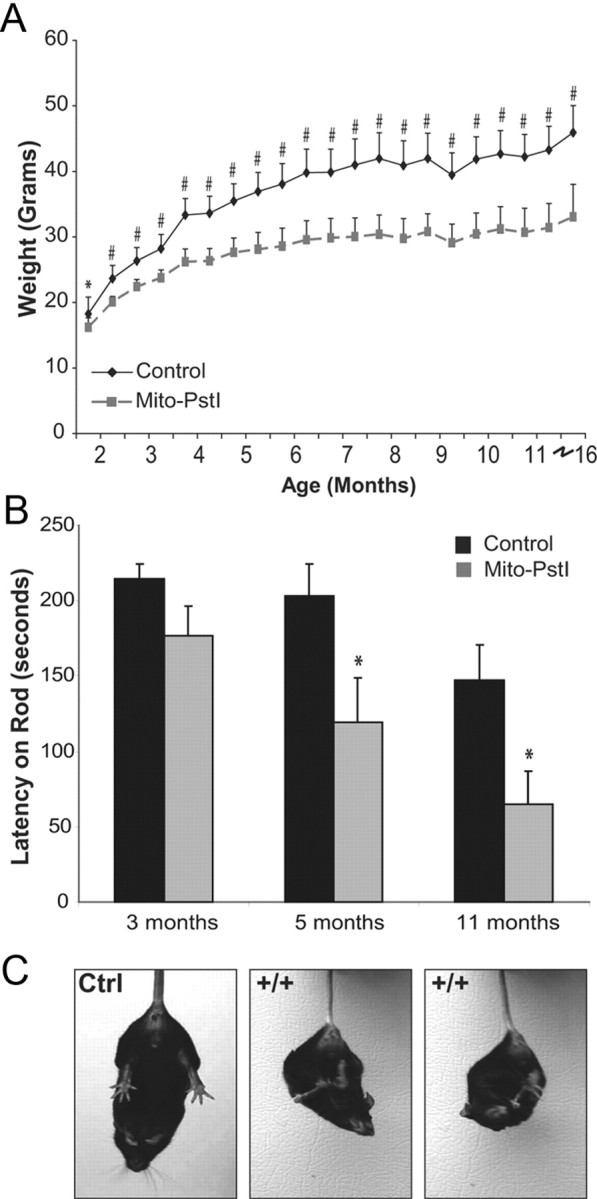
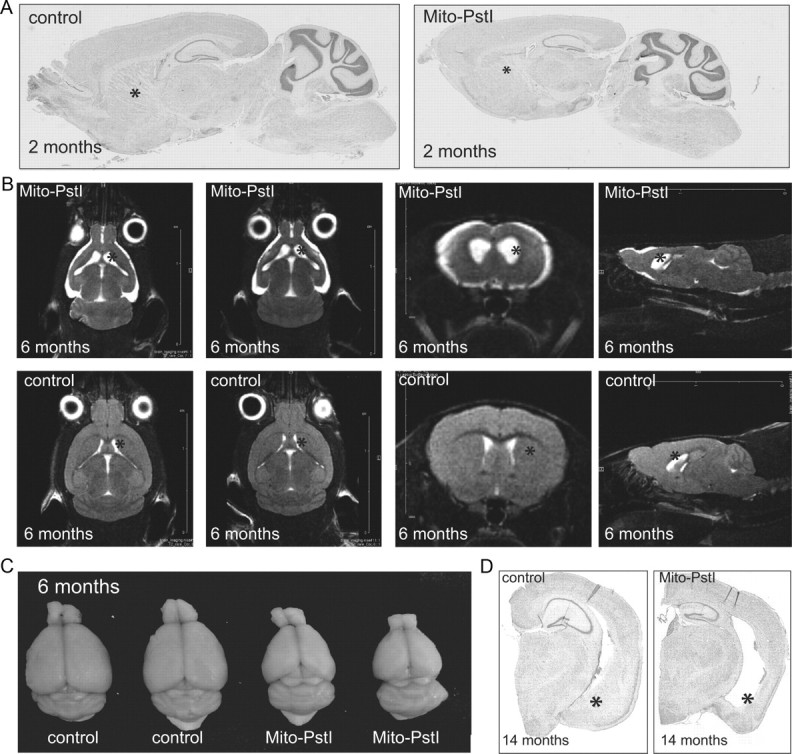
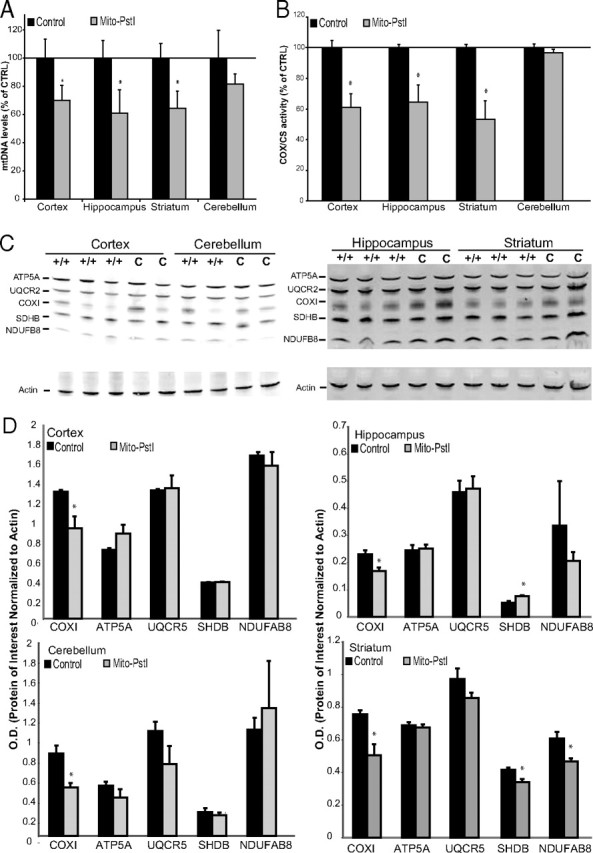
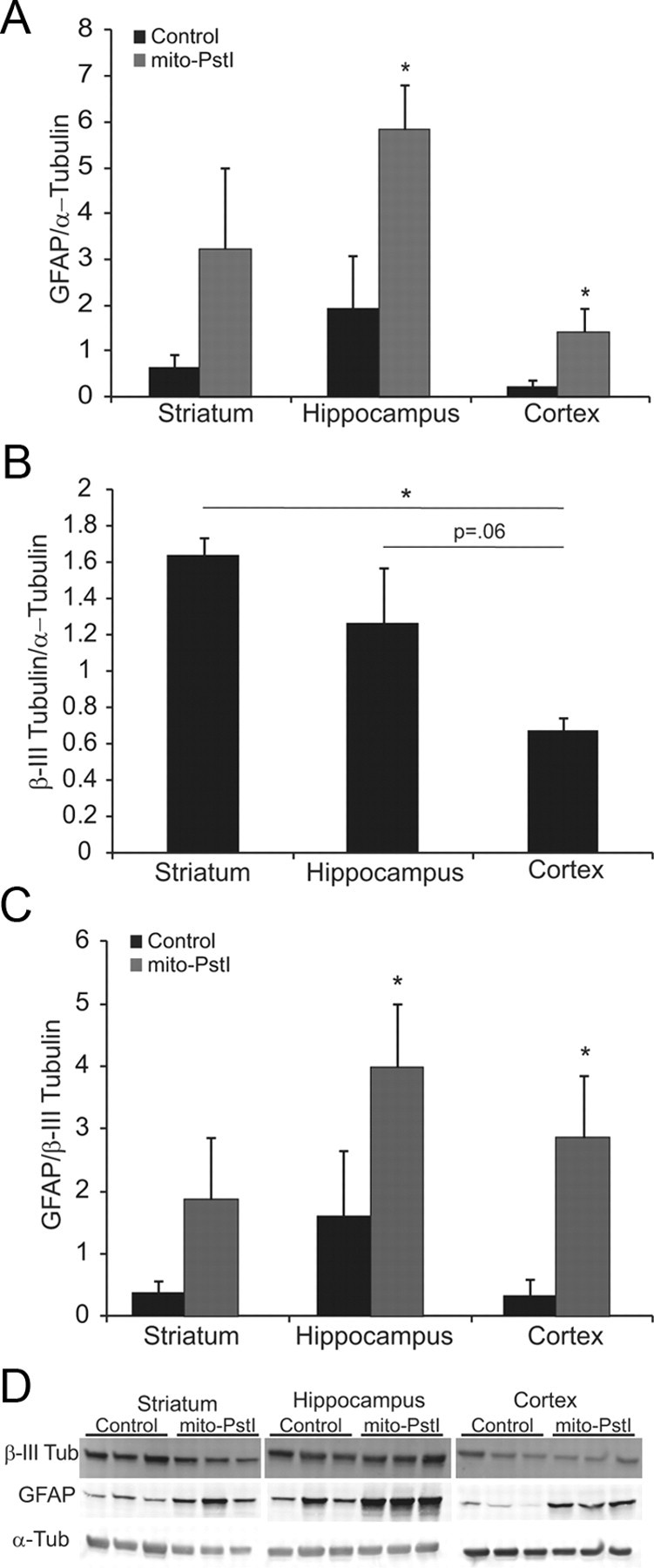
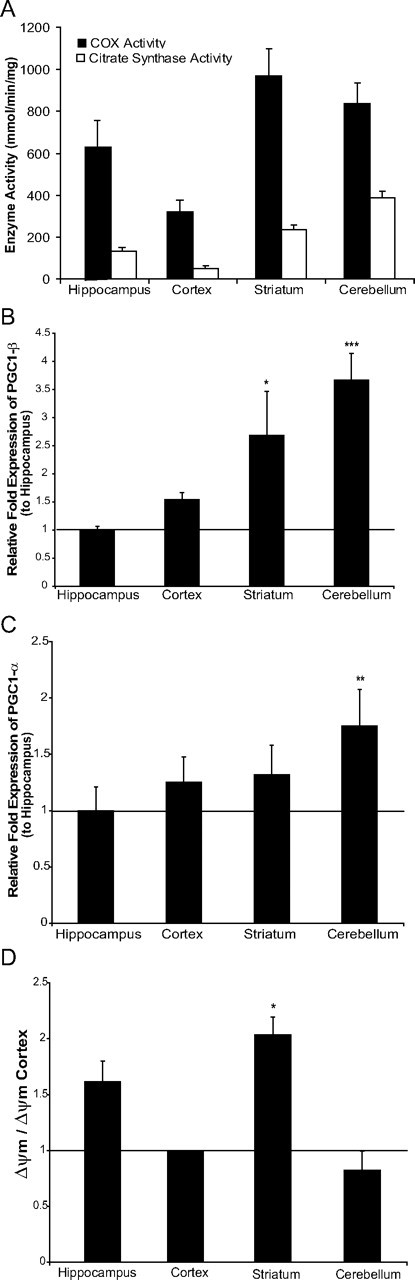
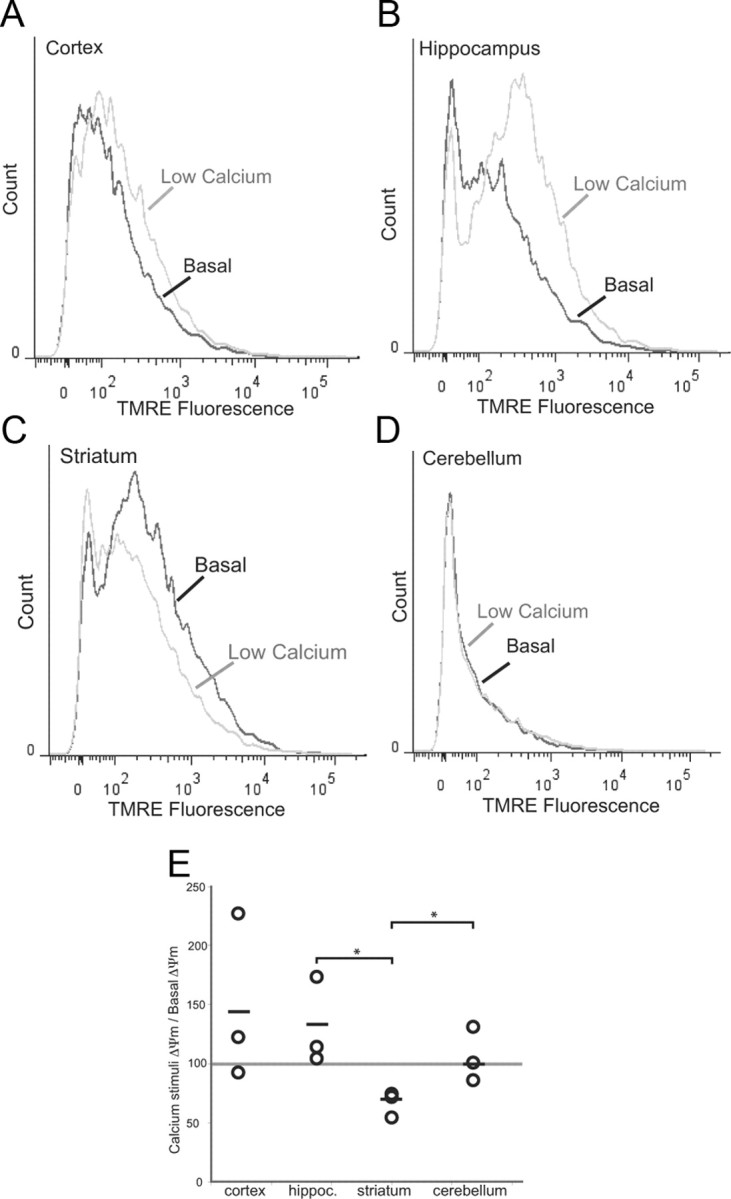
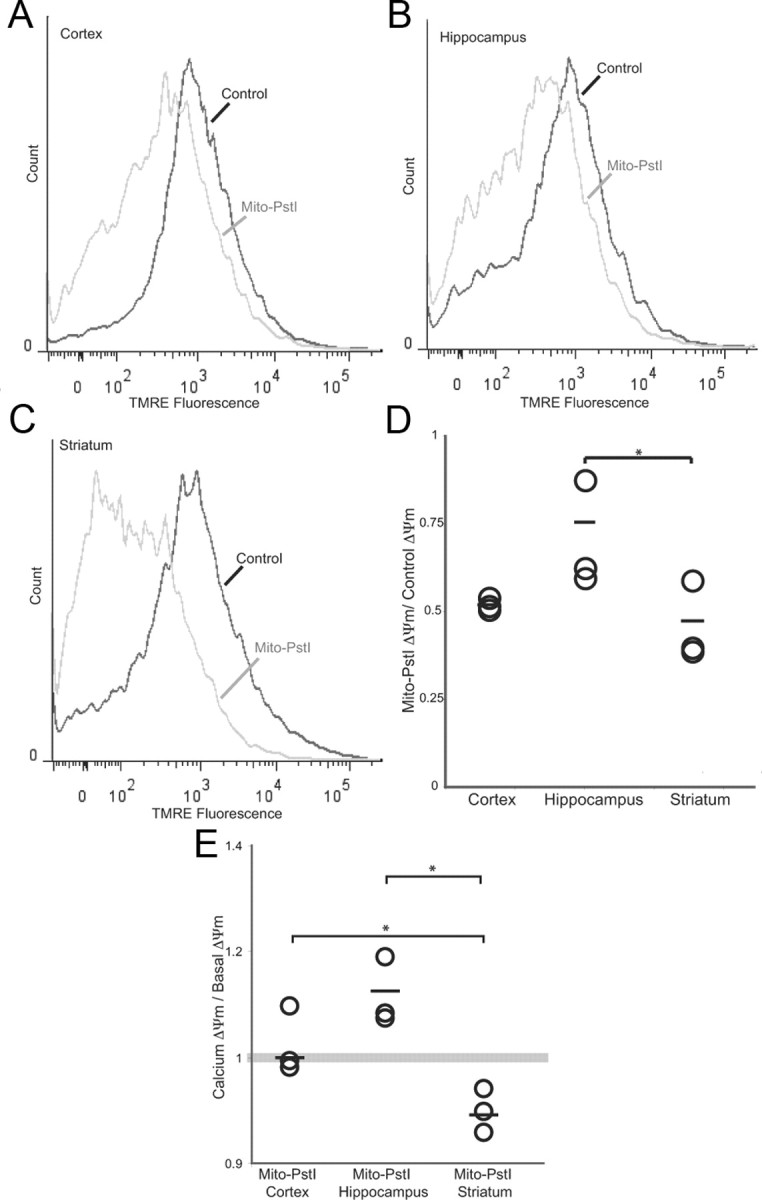
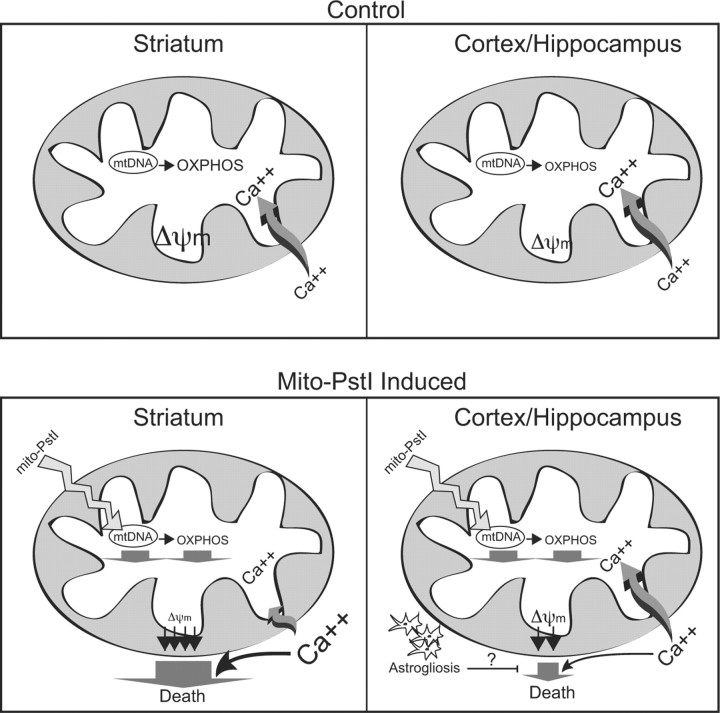
Neuronal oxidative phosphorylation (OXPHOS) deficiency has been associated with a variety of neurodegenerative diseases, including Parkinson's disease and Huntington's disease. However, it is not clear how mitochondrial dysfunction alone can lead to a preferential elimination of certain neuronal populations in vivo. We compared different types of neuronal populations undergoing the same OXPHOS deficiency to determine their relative susceptibility and mechanisms responsible for selective neuron vulnerability. We used a mouse model expressing a mitochondria-targeted restriction enzyme, PstI or mito-PstI. The expression of mito-PstI induces double-strand breaks in the mitochondrial DNA (mtDNA), leading to OXPHOS deficiency, mostly due to mtDNA depletion. We targeted mito-PstI expression to the cortex, hippocampus, and striatum under the CaMKII-α promoter. Animals undergoing long-term expression of mito-PstI displayed a selective worsening of the striatum over cortical and hippocampal areas. Mito-PstI expression and mtDNA depletion were not worse in the striatum, but the latter showed the most severe defects in mitochondrial membrane potential, response to calcium, and survival. These results showed that the striatum is particularly sensitive to defects in OXPHOS possibly due to an increased reliance on OXPHOS function in this area and differences in response to physiological stimuli. These results may help explain the neuropathological features associated with Huntington's disease, which have been associated with OXPHOS defects.
Figures
Similar articles
-
Striatal dysfunctions associated with mitochondrial DNA damage in dopaminergic neurons in a mouse model of Parkinson's disease.J Neurosci. 2011 Nov 30;31(48):17649-58. doi: 10.1523/JNEUROSCI.4871-11.2011. J Neurosci. 2011. PMID: 22131425 Free PMC article.
-
Differential expression of oxidative phosphorylation genes in patients with Alzheimer's disease: implications for early mitochondrial dysfunction and oxidative damage.Neuromolecular Med. 2004;5(2):147-62. doi: 10.1385/NMM:5:2:147. Neuromolecular Med. 2004. PMID: 15075441
-
A stagewise response to mitochondrial dysfunction in mitochondrial DNA maintenance disorders.Biochim Biophys Acta Mol Basis Dis. 2024 Jun;1870(5):167131. doi: 10.1016/j.bbadis.2024.167131. Epub 2024 Mar 21. Biochim Biophys Acta Mol Basis Dis. 2024. PMID: 38521420
-
Significance of Mitochondria DNA Mutations in Diseases.Adv Exp Med Biol. 2017;1038:219-230. doi: 10.1007/978-981-10-6674-0_15. Adv Exp Med Biol. 2017. PMID: 29178079 Review.
-
Different metabolic properties of mitochondrial oxidative phosphorylation in different cell types--important implications for mitochondrial cytopathies.Exp Physiol. 2003 Jan;88(1):149-54. doi: 10.1113/eph8802512. Exp Physiol. 2003. PMID: 12525863 Review.
Cited by
-
Mouse models of Parkinson's disease associated with mitochondrial dysfunction.Mol Cell Neurosci. 2013 Jul;55:87-94. doi: 10.1016/j.mcn.2012.08.002. Epub 2012 Aug 11. Mol Cell Neurosci. 2013. PMID: 22954895 Free PMC article. Review.
-
Mitochondrial mutations and polymorphisms in psychiatric disorders.Front Genet. 2012 Jun 20;3:103. doi: 10.3389/fgene.2012.00103. eCollection 2012. Front Genet. 2012. PMID: 22723804 Free PMC article.
-
Endogenous Parkin Preserves Dopaminergic Substantia Nigral Neurons following Mitochondrial DNA Mutagenic Stress.Neuron. 2015 Jul 15;87(2):371-81. doi: 10.1016/j.neuron.2015.06.034. Neuron. 2015. PMID: 26182419 Free PMC article.
-
Transient systemic mtDNA damage leads to muscle wasting by reducing the satellite cell pool.Hum Mol Genet. 2013 Oct 1;22(19):3976-86. doi: 10.1093/hmg/ddt251. Epub 2013 Jun 10. Hum Mol Genet. 2013. PMID: 23760083 Free PMC article.
-
Adult-Onset Deficiency of Mitochondrial Complex III in a Mouse Model of Alzheimer's Disease Decreases Amyloid Beta Plaque Formation.Mol Neurobiol. 2022 Oct;59(10):6552-6566. doi: 10.1007/s12035-022-02992-3. Epub 2022 Aug 15. Mol Neurobiol. 2022. PMID: 35969330 Free PMC article.
References
-
- Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, Schreier PH, Smith AJ, Staden R, Young IG. Sequence and organization of the human mitochondrial genome. Nature. 1981;290:457–465. - PubMed
-
- Barrientos A. In vivo and in organello assessment of OXPHOS activities. Methods. 2002;26:307–316. - PubMed
-
- Berthet C, Lei H, Thevenet J, Gruetter R, Magistretti PJ, Hirt L. Neuroprotective role of lactate after cerebral ischemia. J Cereb Blood Flow Metab. 2009;29:1780–1789. - PubMed
-
- Betarbet R, Sherer TB, MacKenzie G, Garcia-Osuna M, Panov AV, Greenamyre JT. Chronic systemic pesticide exposure reproduces features of Parkinson's disease. Nat Neurosci. 2000;3:1301–1306. - PubMed
-
- Brookes PS, Yoon Y, Robotham JL, Anders MW, Sheu SS. Calcium, ATP, and ROS: a mitochondrial love-hate triangle. Am J Physiol Cell Physiol. 2004;287:C817–833. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases