

Mosaic analysis with double markers reveals tumor cell of origin in glioma
- PMID: 21737130
- PMCID: PMC3143261
- DOI: 10.1016/j.cell.2011.06.014
Mosaic analysis with double markers reveals tumor cell of origin in glioma
Abstract
Cancer cell of origin is difficult to identify by analyzing cells within terminal stage tumors, whose identity could be concealed by the acquired plasticity. Thus, an ideal approach to identify the cell of origin is to analyze proliferative abnormalities in distinct lineages prior to malignancy. Here, we use mosaic analysis with double markers (MADM) in mice to model gliomagenesis by initiating concurrent p53/Nf1 mutations sporadically in neural stem cells (NSCs). Surprisingly, MADM-based lineage tracing revealed significant aberrant growth prior to malignancy only in oligodendrocyte precursor cells (OPCs), but not in any other NSC-derived lineages or NSCs themselves. Upon tumor formation, phenotypic and transcriptome analyses of tumor cells revealed salient OPC features. Finally, introducing the same p53/Nf1 mutations directly into OPCs consistently led to gliomagenesis. Our findings suggest OPCs as the cell of origin in this model, even when initial mutations occur in NSCs, and highlight the importance of analyzing premalignant stages to identify the cancer cell of origin.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
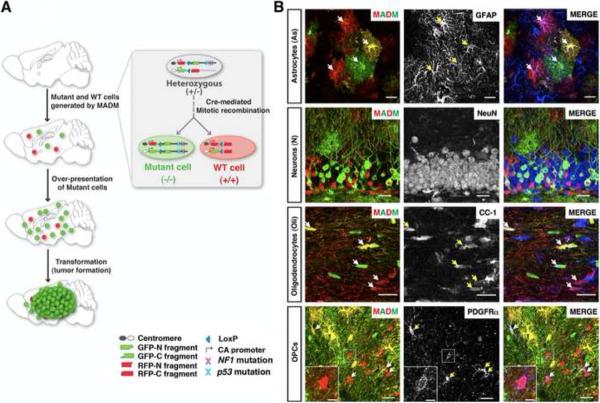
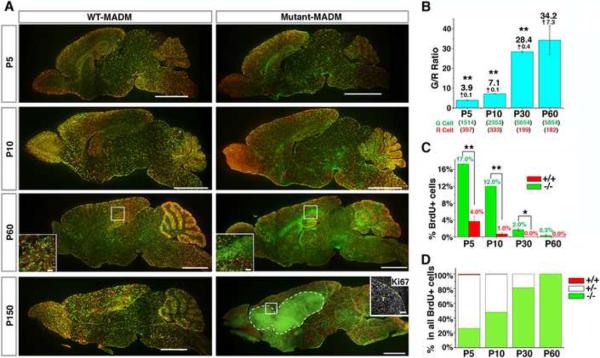
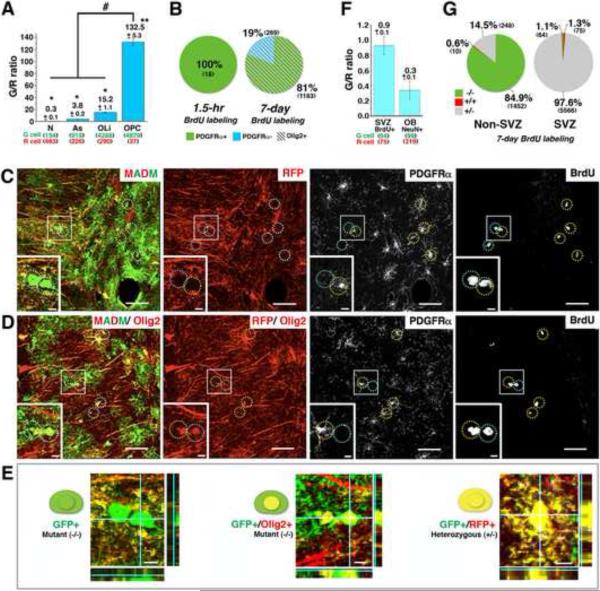
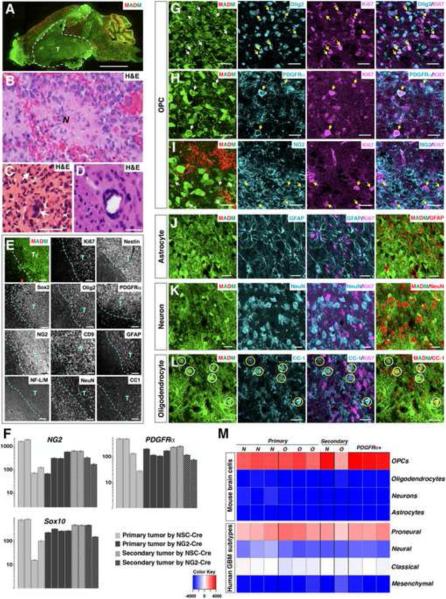
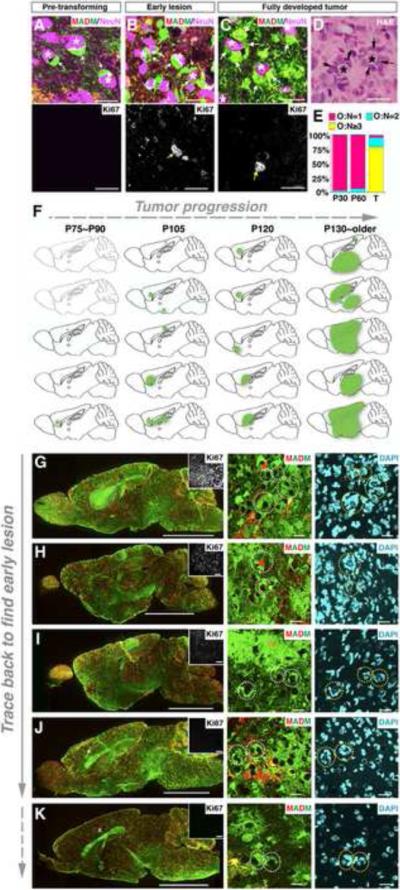
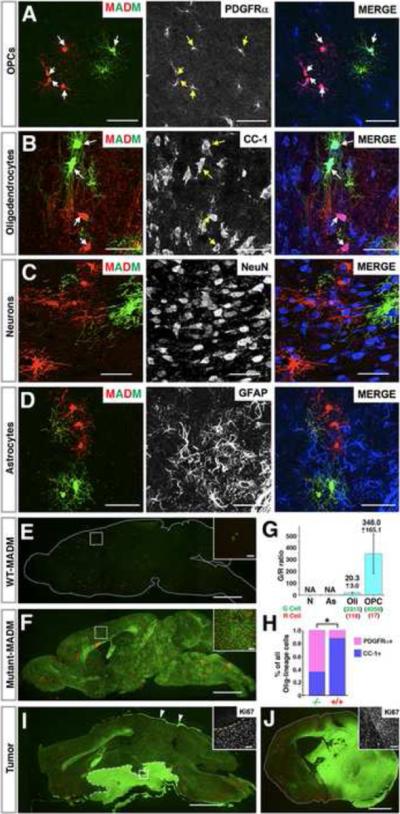
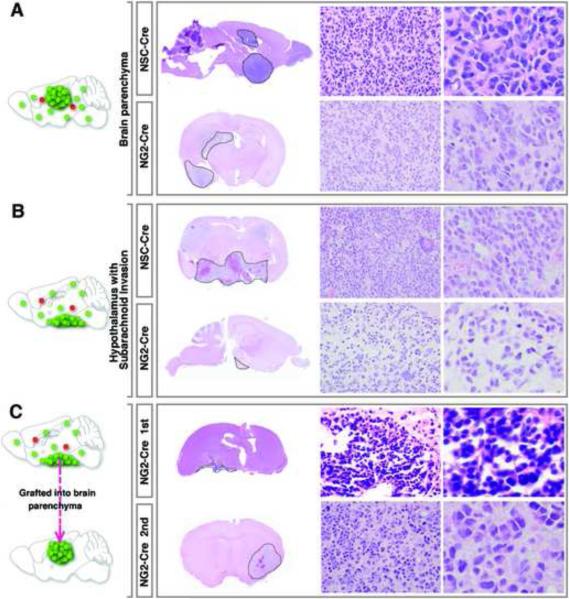
Comment in
-
Glioma development: where did it all go wrong?Cell. 2011 Jul 22;146(2):187-8. doi: 10.1016/j.cell.2011.06.047. Cell. 2011. PMID: 21784240
-
Tumorigenesis: the origins of glioma.Nat Rev Cancer. 2011 Aug 24;11(9):627. doi: 10.1038/nrc3129. Nat Rev Cancer. 2011. PMID: 21863049 No abstract available.
References
-
- Bachoo RM, Maher EA, Ligon KL, Sharpless NE, Chan SS, You MJ, Tang Y, DeFrances J, Stover E, Weissleder R, et al. Epidermal growth factor receptor and Ink4a/Arf: convergent mechanisms governing terminal differentiation and transformation along the neural stem cell to astrocyte axis. Cancer Cell. 2002;1:269–277. - PubMed
-
- Cahoy JD, Emery B, Kaushal A, Foo LC, Zamanian JL, Christopherson KS, Xing Y, Lubischer JL, Krieg PA, Krupenko SA, et al. A transcriptome database for astrocytes, neurons, and oligodendrocytes: a new resource for understanding brain development and function. J Neurosci. 2008;28:264–278. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
