

Regulators of G-protein signaling and their Gα substrates: promises and challenges in their use as drug discovery targets
- PMID: 21737532
- PMCID: PMC3141876
- DOI: 10.1124/pr.110.003038
Regulators of G-protein signaling and their Gα substrates: promises and challenges in their use as drug discovery targets
Abstract
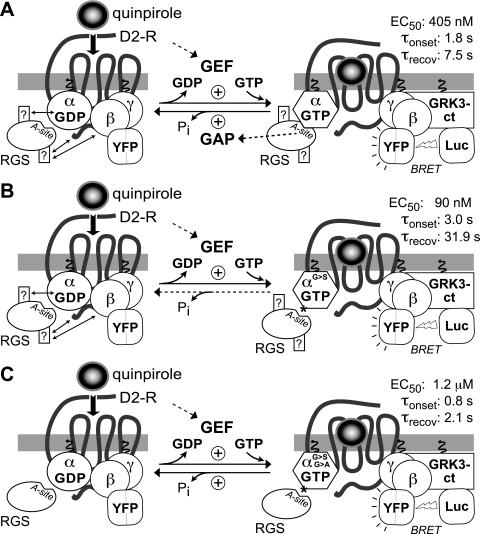
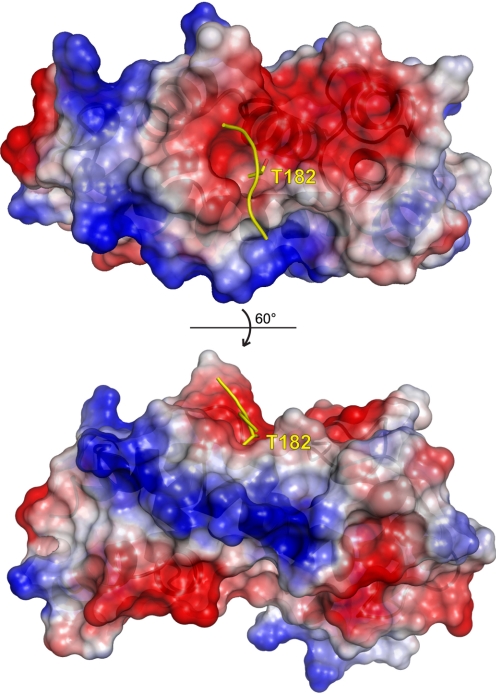
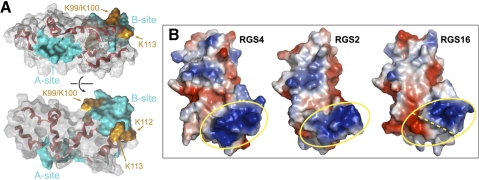
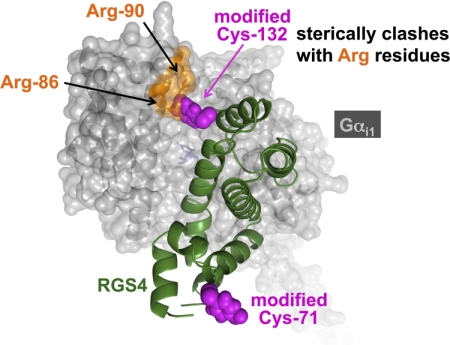
Because G-protein coupled receptors (GPCRs) continue to represent excellent targets for the discovery and development of small-molecule therapeutics, it is posited that additional protein components of the signal transduction pathways emanating from activated GPCRs themselves are attractive as drug discovery targets. This review considers the drug discovery potential of two such components: members of the "regulators of G-protein signaling" (RGS protein) superfamily, as well as their substrates, the heterotrimeric G-protein α subunits. Highlighted are recent advances, stemming from mouse knockout studies and the use of "RGS-insensitivity" and fast-hydrolysis mutations to Gα, in our understanding of how RGS proteins selectively act in (patho)physiologic conditions controlled by GPCR signaling and how they act on the nucleotide cycling of heterotrimeric G-proteins in shaping the kinetics and sensitivity of GPCR signaling. Progress is documented regarding recent activities along the path to devising screening assays and chemical probes for the RGS protein target, not only in pursuits of inhibitors of RGS domain-mediated acceleration of Gα GTP hydrolysis but also to embrace the potential of finding allosteric activators of this RGS protein action. The review concludes in considering the Gα subunit itself as a drug target, as brought to focus by recent reports of activating mutations to GNAQ and GNA11 in ocular (uveal) melanoma. We consider the likelihood of several strategies for antagonizing the function of these oncogene alleles and their gene products, including the use of RGS proteins with Gα(q) selectivity.
Figures
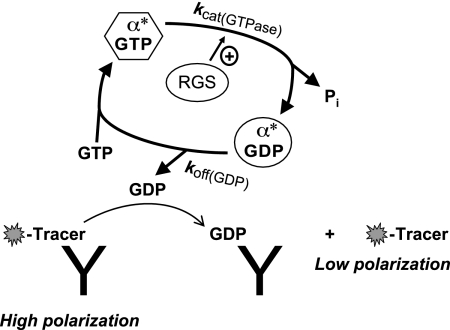
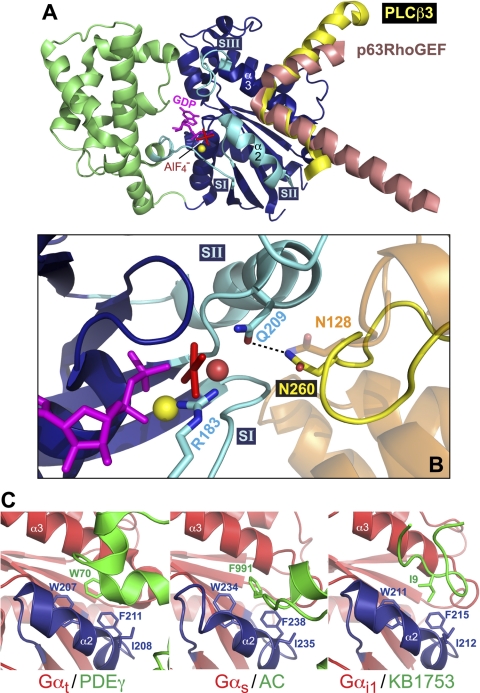
References
-
- Anger T, Zhang W, Mende U. (2004) Differential contribution of GTPase activation and effector antagonism to the inhibitory effect of RGS proteins on Gq-mediated signaling in vivo. J Biol Chem 279:3906–3915 - PubMed
-
- Apanovitch DM, Slep KC, Sigler PB, Dohlman HG. (1998) Sst2 is a GTPase-activating protein for Gpa1: purification and characterization of a cognate RGS-Galpha protein pair in yeast. Biochemistry 37:4815–4822 - PubMed
-
- Arshavsky VY, Lamb TD, Pugh EN., Jr (2002) G proteins and phototransduction. Annu Rev Physiol 64:153–187 - PubMed
-
- Ashihara E, Kawata E, Maekawa T. (2010) Future prospect of RNA interference for cancer therapies. Curr Drug Targets 11:345–360 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01-GM074268/GM/NIGMS NIH HHS/United States
- F30 MH074266/MH/NIMH NIH HHS/United States
- R01 GM062338/GM/NIGMS NIH HHS/United States
- R01-GM062338/GM/NIGMS NIH HHS/United States
- R01 GM074268/GM/NIGMS NIH HHS/United States
- T32-GM007040/GM/NIGMS NIH HHS/United States
- R03 DA030555/DA/NIDA NIH HHS/United States
- R03-DA030555/DA/NIDA NIH HHS/United States
- R01-GM082892/GM/NIGMS NIH HHS/United States
- R01 GM082892/GM/NIGMS NIH HHS/United States
- T32 GM007040/GM/NIGMS NIH HHS/United States
- F30-MH074266/MH/NIMH NIH HHS/United States
- T32-GM008719/GM/NIGMS NIH HHS/United States
- T32 GM008719/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
