

RNA transcript 3'-proximal sequence affects translocation bias of RNA polymerase
- PMID: 21739957
- PMCID: PMC3322512
- DOI: 10.1021/bi200437q
RNA transcript 3'-proximal sequence affects translocation bias of RNA polymerase
Abstract
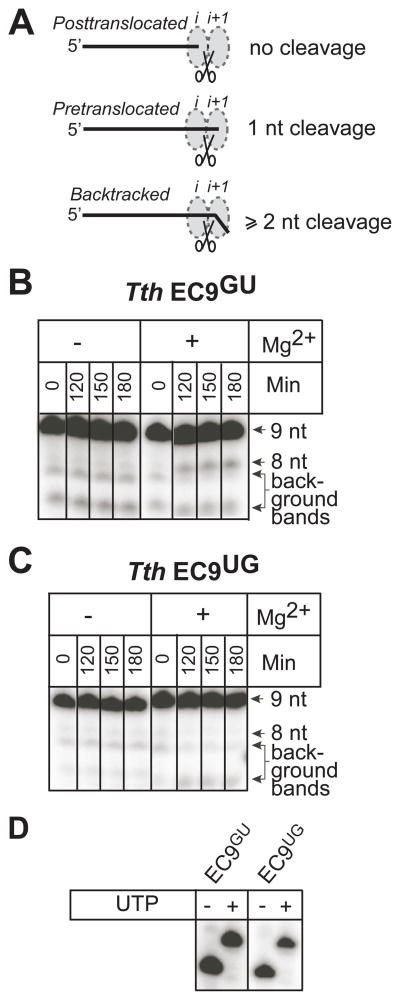
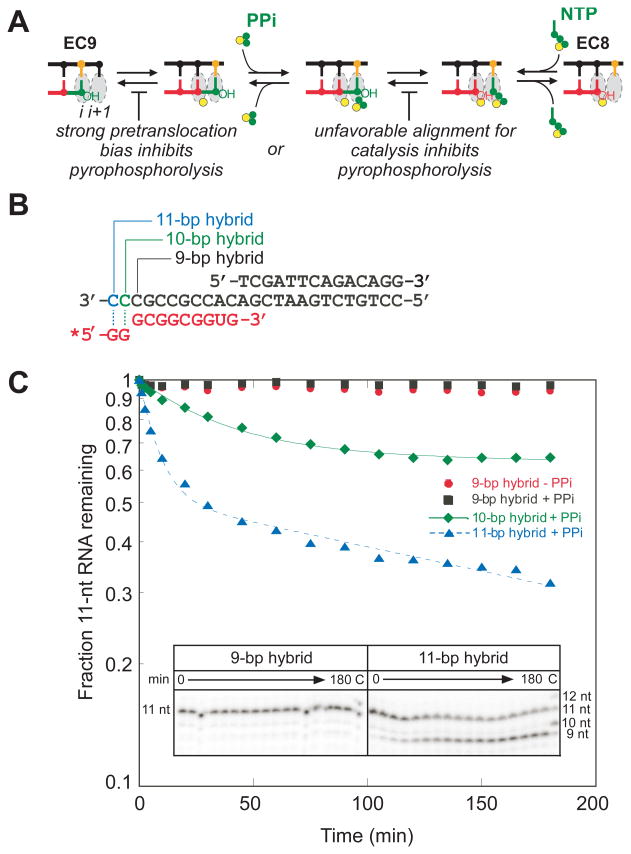
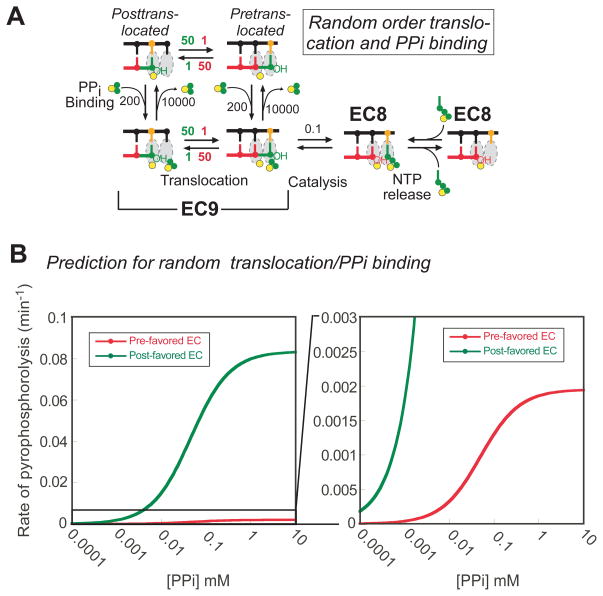
Translocation of RNA polymerase on DNA is thought to involve oscillations between pretranslocated and posttranslocated states that are rectified by nucleotide addition or pyrophosphorolysis. The pretranslocated register is also a precursor to transcriptional pause states that mediate regulation of transcript elongation. However, the determinants of bias between the pretranslocated and posttranslocated states are incompletely understood. To investigate translocation bias in multisubunit RNA polymerases, we measured rates of pyrophosphorolysis, which occurs in the pretranslocated register, in minimal elongation complexes containing T. thermophilus or E. coli RNA polymerase. Our results suggest that the identity of RNA:DNA nucleotides in the active site are strong determinants of susceptibility to pyrophosphorolysis, and thus translocation bias, with the 3' RNA nucleotide favoring the pretranslocated state in the order U > C > A > G. The preference of 3' U vs G for the pretranslocated register appeared to be universal among both bacterial and eukaryotic RNA polymerases and was confirmed by exonuclease III footprinting of defined elongation complexes. However, the relationship of pyrophosphate concentration to the rate of pyrophosphorolysis of 3' U-containing versus 3' G-containing elongation complexes did not match predictions of a simple mechanism in which 3'-RNA seqeunce affects only translocation bias and pyrophosphate (PPi) binds only to the pretranslocated state.
Figures






References
-
- Zhang JaLR. Substrate loading, nucleotide addition, and translocation by RNA Polymerase. In: Buc HaST., editor. RNA Polymerase as Molecular Motors. 1. Royal Society of Chemistry; Cambridge, UK: 2009. pp. 206–235.
-
- Kireeva M, Kashlev M, Burton ZF. Translocation by multi-subunit RNA polymerases. Biochim Biophys Acta 2010 - PubMed
-
- Guajardo R, Sousa R. A model for the mechanism of polymerase translocation. J Mol Biol. 1997;265:8–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
