

Interferon regulatory factor-1 binds c-Cbl, enhances mitogen activated protein kinase signaling and promotes retinoic acid-induced differentiation of HL-60 human myelo-monoblastic leukemia cells
- PMID: 21740303
- PMCID: PMC3989140
- DOI: 10.3109/10428194.2011.603449
Interferon regulatory factor-1 binds c-Cbl, enhances mitogen activated protein kinase signaling and promotes retinoic acid-induced differentiation of HL-60 human myelo-monoblastic leukemia cells
Abstract
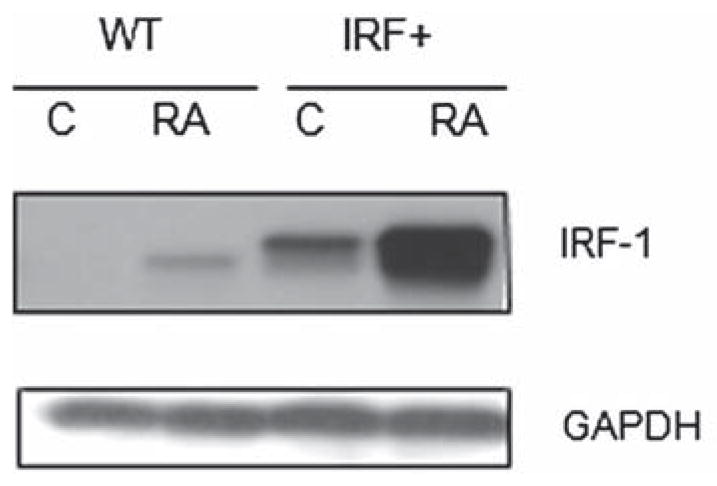
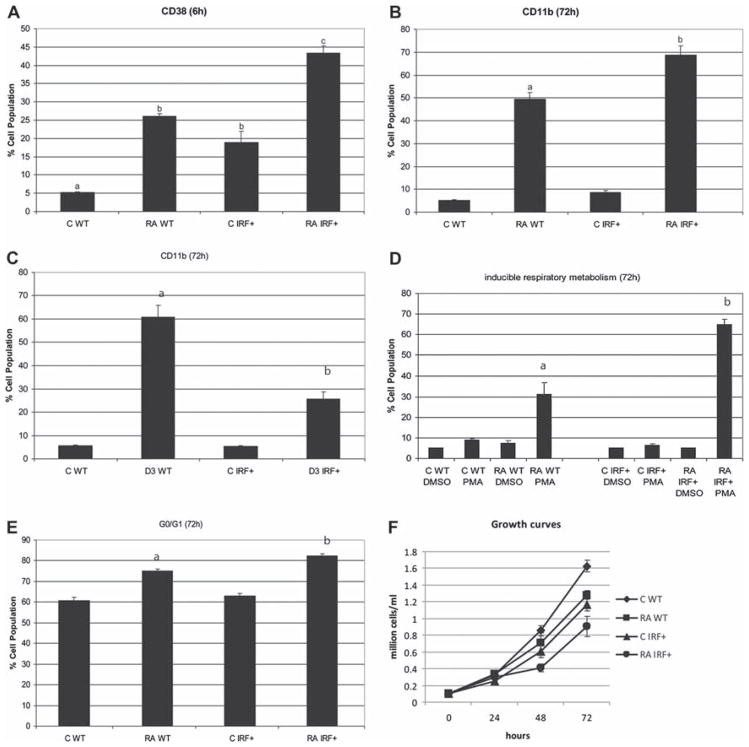
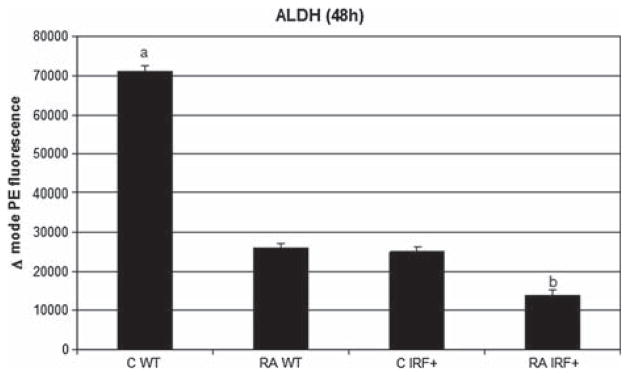
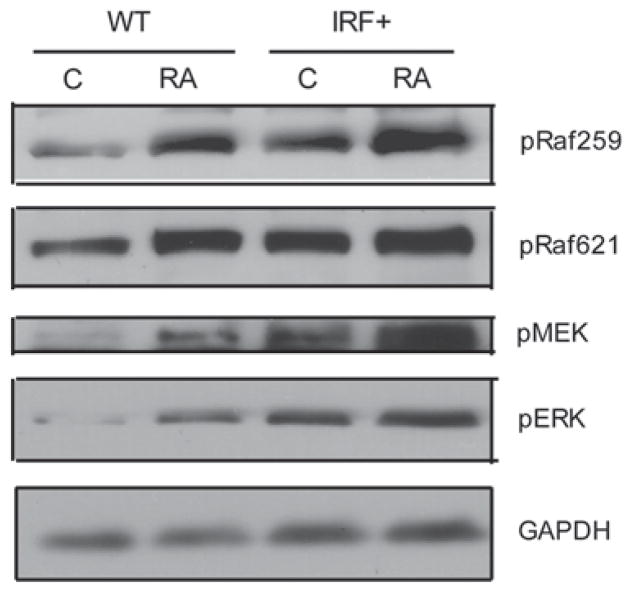
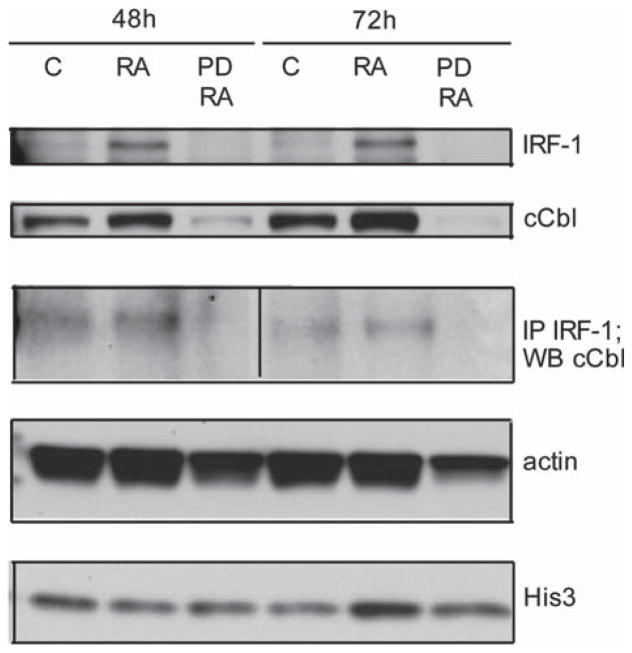
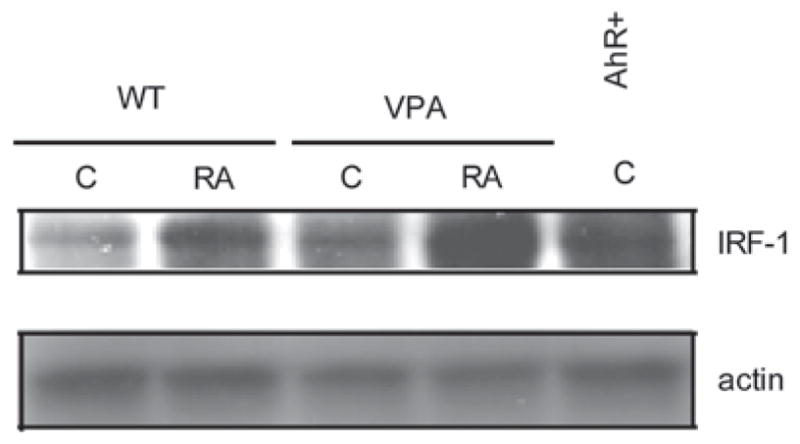
All-trans retinoic acid (RA) and interferons (IFNs) have efficacy in treating certain leukemias and lymphomas, respectively, motivating interest in their mechanism of action to improve therapy. Both RA and IFNs induce interferon regulatory factor-1 (IRF-1). We find that in HL-60 myeloblastic leukemia cells which undergo mitogen activated protien kinase (MAPK)-dependent myeloid differentiation in response to RA, IRF-1 propels differentiation. RA induces MAPK-dependent expression of IRF-1. IRF-1 binds c-Cbl, a MAPK related adaptor. Ectopic IRF-1 expression causes CD38 expression and activation of the Raf/MEK/ERK axis, and enhances RA-induced differentiation by augmenting CD38, CD11b, respiratory burst and G0 arrest. Ectopic IRF-1 expression also decreases the activity of aldehyde dehydrogenase 1, a stem cell marker, and enhances RA-induced ALDH1 down-regulation. Interestingly, expression of aryl hydrocarbon receptor (AhR), which is RA-induced and known to down-regulate Oct4 and drive RA-induced differentiation, also enhances IRF-1 expression. The data are consistent with a model whereby IRF-1 acts downstream of RA and AhR to enhance Raf/MEK/ERK activation and propel differentiation.
Conflict of interest statement
Figures
Similar articles
-
6-Formylindolo(3,2-b)Carbazole (FICZ) Modulates the Signalsome Responsible for RA-Induced Differentiation of HL-60 Myeloblastic Leukemia Cells.PLoS One. 2015 Aug 19;10(8):e0135668. doi: 10.1371/journal.pone.0135668. eCollection 2015. PLoS One. 2015. PMID: 26287494 Free PMC article.
-
c-Cbl tyrosine kinase-binding domain mutant G306E abolishes the interaction of c-Cbl with CD38 and fails to promote retinoic acid-induced cell differentiation and G0 arrest.J Biol Chem. 2009 Sep 18;284(38):25664-77. doi: 10.1074/jbc.M109.014241. Epub 2009 Jul 27. J Biol Chem. 2009. PMID: 19635790 Free PMC article.
-
6-Formylindolo (3,2-b)carbazole (FICZ) enhances retinoic acid (RA)-induced differentiation of HL-60 myeloblastic leukemia cells.Mol Cancer. 2013 May 9;12:39. doi: 10.1186/1476-4598-12-39. Mol Cancer. 2013. PMID: 23656719 Free PMC article.
-
Activation of the aryl hydrocarbon receptor AhR Promotes retinoic acid-induced differentiation of myeloblastic leukemia cells by restricting expression of the stem cell transcription factor Oct4.Cancer Res. 2011 Mar 15;71(6):2371-80. doi: 10.1158/0008-5472.CAN-10-2299. Epub 2011 Jan 24. Cancer Res. 2011. PMID: 21262915 Free PMC article.
-
Retinoic acid and interferon signaling cross talk in normal and RA-resistant APL cells.Leukemia. 1999 Aug;13(8):1167-74. doi: 10.1038/sj.leu.2401469. Leukemia. 1999. PMID: 10450744 Review.
Cited by
-
Induced myelomonocytic differentiation in leukemia cells is accompanied by noncanonical transcription factor expression.FEBS Open Bio. 2015 Sep 28;5:789-800. doi: 10.1016/j.fob.2015.09.008. eCollection 2015. FEBS Open Bio. 2015. PMID: 26566473 Free PMC article.
-
6-Formylindolo(3,2-b)Carbazole (FICZ) Modulates the Signalsome Responsible for RA-Induced Differentiation of HL-60 Myeloblastic Leukemia Cells.PLoS One. 2015 Aug 19;10(8):e0135668. doi: 10.1371/journal.pone.0135668. eCollection 2015. PLoS One. 2015. PMID: 26287494 Free PMC article.
-
Isocitrate dehydrogenase 1 mutations prime the all-trans retinoic acid myeloid differentiation pathway in acute myeloid leukemia.J Exp Med. 2016 Apr 4;213(4):483-97. doi: 10.1084/jem.20150736. Epub 2016 Mar 7. J Exp Med. 2016. PMID: 26951332 Free PMC article. Clinical Trial.
-
An integrated approach to elucidate signaling pathways of dioscin-induced apoptosis, energy metabolism and differentiation in acute myeloid leukemia.Naunyn Schmiedebergs Arch Pharmacol. 2018 Jun;391(6):587-602. doi: 10.1007/s00210-018-1484-6. Epub 2018 Mar 28. Naunyn Schmiedebergs Arch Pharmacol. 2018. PMID: 29594316
-
An Effective Model of the Retinoic Acid Induced HL-60 Differentiation Program.Sci Rep. 2017 Oct 30;7(1):14327. doi: 10.1038/s41598-017-14523-5. Sci Rep. 2017. PMID: 29085021 Free PMC article.
References
-
- Tefferi A, Mathew P, Noel P. The 5q-syndrome: a scientific and clinical update. Leuk Lymphoma. 1994;14:375–378. - PubMed
-
- Willman CL, Sever CE, Pallavicini MG, et al. Deletion of IRF-1, mapping to chromosome 5q31. 1, in human leukemia and preleukemic myelodysplasia. Science. 1993;259:968–971. - PubMed
-
- Harada H, Kondo T, Ogawa S, et al. Accelerated exon skipping of IRF-1 mRNA in human myelodysplasia/leukemia; a possible mechanism of tumor suppressor inactivation. Oncogene. 1994;9:3313–3320. - PubMed
-
- Harada H, Willison K, Sakakibara J, et al. Absence of the type I IFN system in EC cells: transcriptional activator (IRF-1) and repressor (IRF-2) genes are developmentally regulated. Cell. 1990;63:303–312. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous