

Evolution of the Alx homeobox gene family: parallel retention and independent loss of the vertebrate Alx3 gene
- PMID: 21740507
- PMCID: PMC3166657
- DOI: 10.1111/j.1525-142X.2011.00489.x
Evolution of the Alx homeobox gene family: parallel retention and independent loss of the vertebrate Alx3 gene
Abstract
The Alx gene family is implicated in craniofacial development and comprises two to four homeobox genes in each vertebrate genome analyzed. Using phylogenetics and comparative genomics, we show that the common ancestor of jawed vertebrates had three Alx genes descendent from the two-round genome duplications (Alx1, Alx3, Alx4), compared with a single amphioxus gene. Later in evolution one of the paralogues, Alx3, was lost independently from at least three different vertebrate lineages, whereas Alx1 and Alx4 were consistently retained. Comparison of spatial gene expression patterns reveals that the three mouse genes have equivalent craniofacial expression to the two chick and frog genes, suggesting that redundancy compensated for gene loss. We suggest that multiple independent loss of one Alx gene was predisposed by extensive and persistent overlap in gene expression between Alx paralogues. Even so, it is unclear whether it was coincidence or evolutionary bias that resulted in the same Alx gene being lost on each occasion, rather than different members of the gene family.
© 2011 Wiley Periodicals, Inc.
Figures
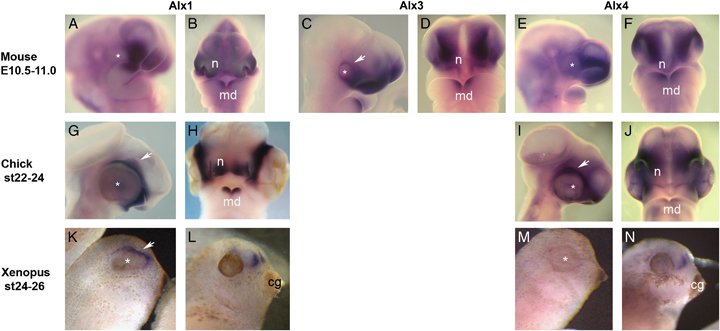
 ). At stages 22–24 of chick development, Alx1 (G, H) and Alx4 (I, J) are also expressed in the facial prominences, again with strong expression around the nasal region (n), and at the distal tip of the mandible (md). In the chick, however, both Alx1 (G) and Alx4 (I) are expressed in the periocular mesenchyme (white arrows). Periocular expression of Alx1 is observed in frog embryos at the stage of 24 (K, white arrow). At this stage, Alx4 is not expressed in frog embryos (M) but later at stage 35, both Alx1 (L) and Alx4 (N) are expressed in frontal mesenchyme above the cement gland (cg).
). At stages 22–24 of chick development, Alx1 (G, H) and Alx4 (I, J) are also expressed in the facial prominences, again with strong expression around the nasal region (n), and at the distal tip of the mandible (md). In the chick, however, both Alx1 (G) and Alx4 (I) are expressed in the periocular mesenchyme (white arrows). Periocular expression of Alx1 is observed in frog embryos at the stage of 24 (K, white arrow). At this stage, Alx4 is not expressed in frog embryos (M) but later at stage 35, both Alx1 (L) and Alx4 (N) are expressed in frontal mesenchyme above the cement gland (cg).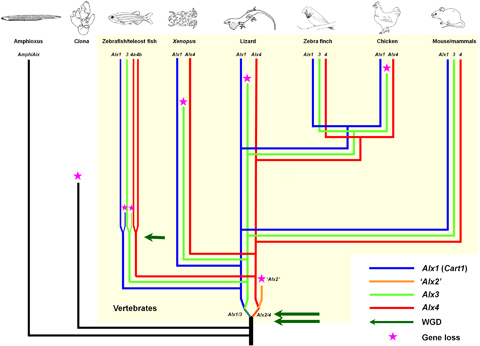
Similar articles
-
Whole genome duplications and expansion of the vertebrate GATA transcription factor gene family.BMC Evol Biol. 2009 Aug 20;9:207. doi: 10.1186/1471-2148-9-207. BMC Evol Biol. 2009. PMID: 19695090 Free PMC article.
-
Alx-4: cDNA cloning and characterization of a novel paired-type homeodomain protein.Gene. 1997 Dec 12;203(2):217-23. doi: 10.1016/s0378-1119(97)00497-6. Gene. 1997. PMID: 9426253
-
Alx1, a member of the Cart1/Alx3/Alx4 subfamily of Paired-class homeodomain proteins, is an essential component of the gene network controlling skeletogenic fate specification in the sea urchin embryo.Development. 2003 Jul;130(13):2917-28. doi: 10.1242/dev.00511. Development. 2003. PMID: 12756175
-
The Dlx genes as clues to vertebrate genomics and craniofacial evolution.Semin Cell Dev Biol. 2013 Feb;24(2):110-8. doi: 10.1016/j.semcdb.2012.12.010. Epub 2013 Jan 3. Semin Cell Dev Biol. 2013. PMID: 23291259 Review.
-
Iroquois genes: genomic organization and function in vertebrate neural development.Curr Opin Genet Dev. 2002 Aug;12(4):403-8. doi: 10.1016/s0959-437x(02)00317-9. Curr Opin Genet Dev. 2002. PMID: 12100884 Review.
Cited by
-
ALX1-related frontonasal dysplasia results from defective neural crest cell development and migration.EMBO Mol Med. 2020 Oct 7;12(10):e12013. doi: 10.15252/emmm.202012013. Epub 2020 Sep 11. EMBO Mol Med. 2020. PMID: 32914578 Free PMC article.
-
Differential expression of aristaless-like homeobox 4: a potential marker for gastric adenocarcinoma.Gastroenterol Hepatol Bed Bench. 2016 Fall;9(4):286-294. Gastroenterol Hepatol Bed Bench. 2016. PMID: 27895854 Free PMC article.
-
Fgf8 dosage determines midfacial integration and polarity within the nasal and optic capsules.Dev Biol. 2013 Feb 1;374(1):185-97. doi: 10.1016/j.ydbio.2012.11.014. Epub 2012 Nov 29. Dev Biol. 2013. PMID: 23201021 Free PMC article.
-
Linking Vertebrate Gene Duplications to the New Head Hypothesis.Biology (Basel). 2023 Sep 6;12(9):1213. doi: 10.3390/biology12091213. Biology (Basel). 2023. PMID: 37759612 Free PMC article. Review.
-
AP-2α and AP-2β cooperatively orchestrate homeobox gene expression during branchial arch patterning.Development. 2018 Jan 25;145(2):dev157438. doi: 10.1242/dev.157438. Development. 2018. PMID: 29229773 Free PMC article.
References
-
- Abascal F, Zardoya R, Posada D. ProtTest: selection of best-fit models of protein evolution. Bioinformatics. 2005;21:2104–2105. - PubMed
-
- Beverdam A, Brouwer A, Reijnen M, Korving J, Meijlink F. Severe nasal clefting and abnormal embryonic apoptosis in Alx3/Alx4 double mutant mice. Development. 2001;128:3975–3986. - PubMed
-
- Beverdam A, Meijlink F. Expression patterns of group-I aristaless-related genes during craniofacial and limb development. Mech. Dev. 2001;107:163–167. - PubMed
-
- Bürglin T. A comprehensive classification of homeobox genes. In: Duboule D, editor. Guidebook to the Homeobox Genes. Oxford: Oxford University Press; 1994. pp. 25–73.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
