

Sensory integration for reaching: models of optimality in the context of behavior and the underlying neural circuits
- PMID: 21741553
- PMCID: PMC3361512
- DOI: 10.1016/B978-0-444-53752-2.00004-7
Sensory integration for reaching: models of optimality in the context of behavior and the underlying neural circuits
Abstract
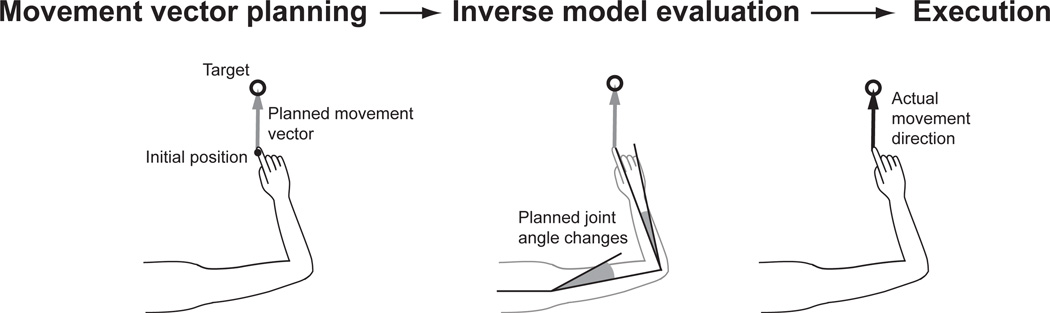
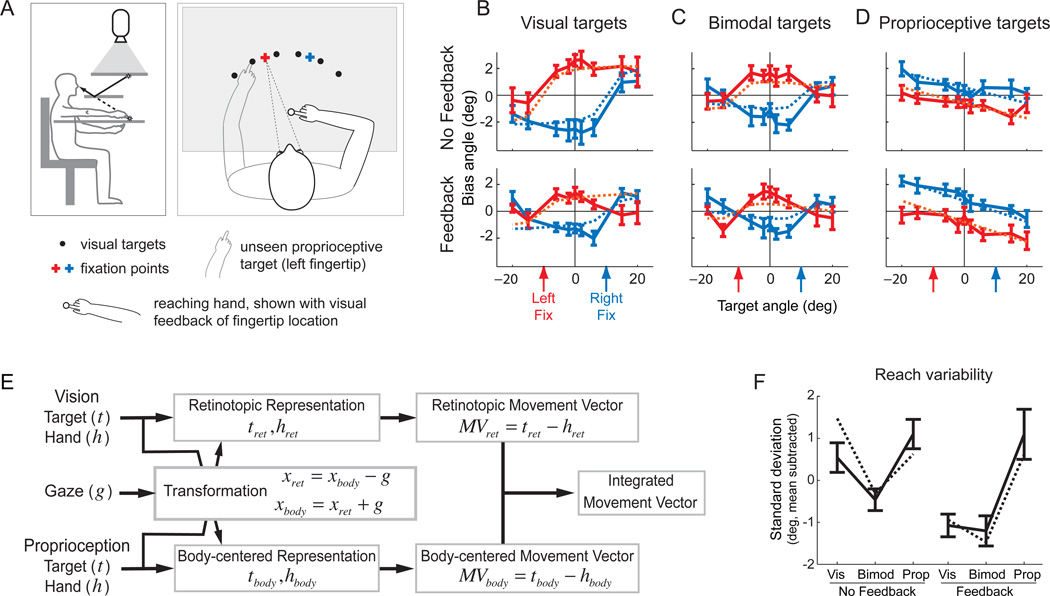
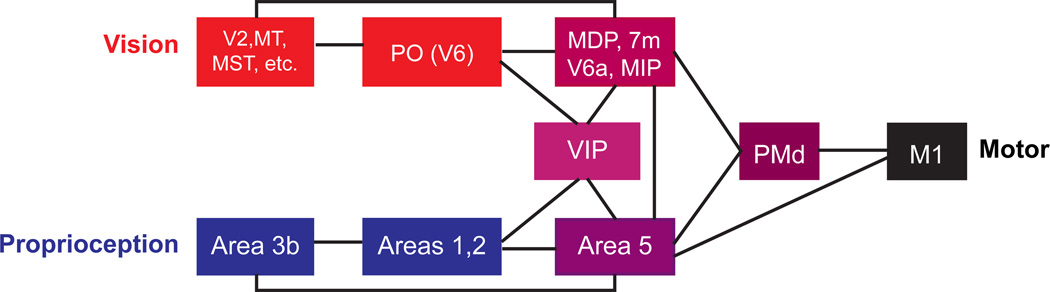
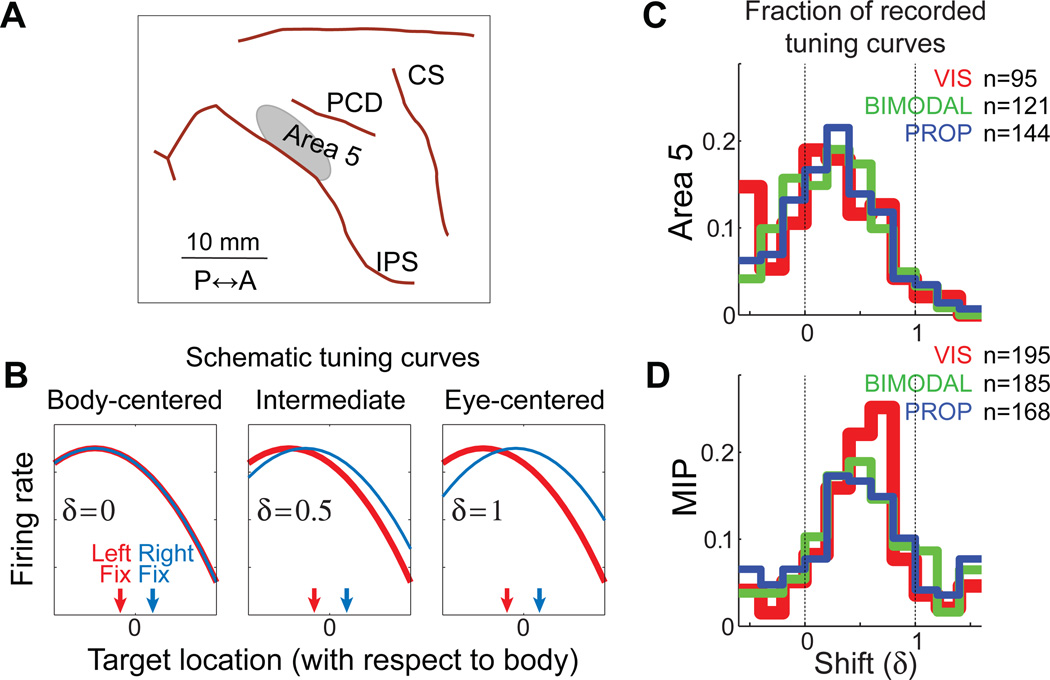
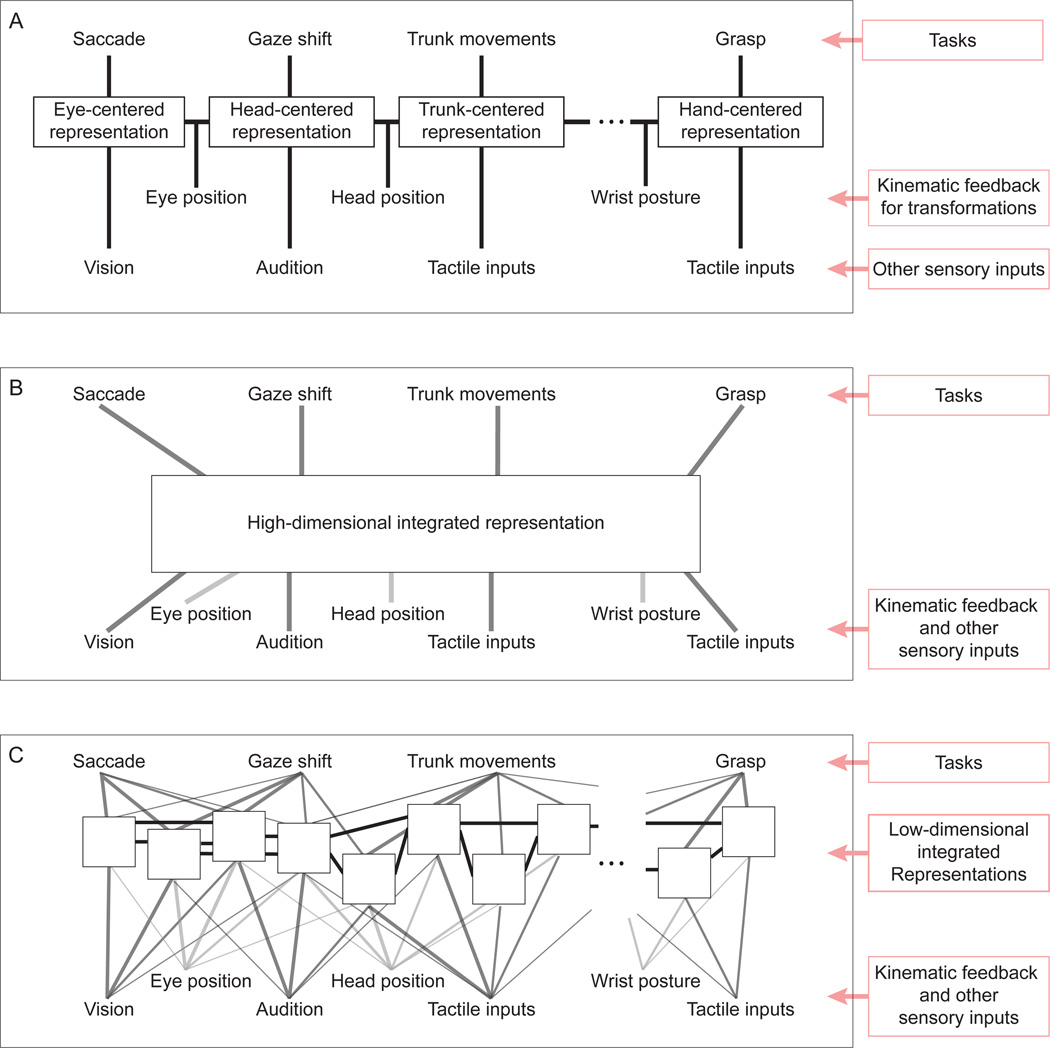
Although multisensory integration has been well modeled at the behavioral level, the link between these behavioral models and the underlying neural circuits is still not clear. This gap is even greater for the problem of sensory integration during movement planning and execution. The difficulty lies in applying simple models of sensory integration to the complex computations that are required for movement control and to the large networks of brain areas that perform these computations. Here I review psychophysical, computational, and physiological work on multisensory integration during movement planning, with an emphasis on goal-directed reaching. I argue that sensory transformations must play a central role in any modeling effort. In particular, the statistical properties of these transformations factor heavily into the way in which downstream signals are combined. As a result, our models of optimal integration are only expected to apply "locally," that is, independently for each brain area. I suggest that local optimality can be reconciled with globally optimal behavior if one views the collection of parietal sensorimotor areas not as a set of task-specific domains, but rather as a palette of complex, sensorimotor representations that are flexibly combined to drive downstream activity and behavior.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures
References
-
- Andersen RA, Buneo CA. Intentional maps in posterior parietal cortex. Annu Rev Neurosci. 2002:189–220. - PubMed
-
- Avillac M, Deneve S, Olivier E, Pouget A, Duhamel JR. Reference frames for representing visual and tactile locations in parietal cortex. Nat Neurosci. 2005;8:941–949. - PubMed
-
- Batista AP, Buneo CA, Snyder LH, Andersen RA. Reach plans in eye-centered coordinates. Science. 1999;285:257–260. - PubMed
-
- Beurze SM, de Lange FP, Toni I, Medendorp WP. Integration of target and effector information in the human brain during reach planning. J Neurophysiol. 2007;97:188–199. - PubMed
-
- Bock O. Localization of objects in the peripheral visual field. Behav Brain Res. 1993;56:77–84. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
