

The FRD3 citrate effluxer promotes iron nutrition between symplastically disconnected tissues throughout Arabidopsis development
- PMID: 21742986
- PMCID: PMC3226209
- DOI: 10.1105/tpc.111.088088
The FRD3 citrate effluxer promotes iron nutrition between symplastically disconnected tissues throughout Arabidopsis development
Abstract
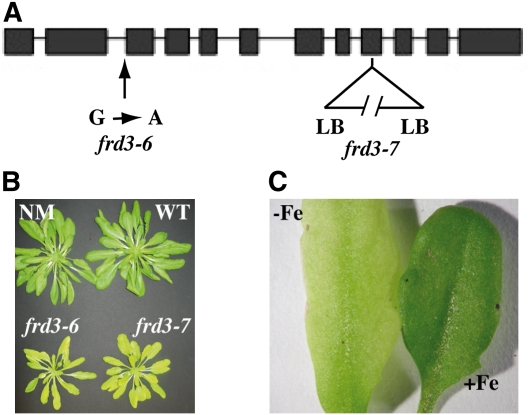
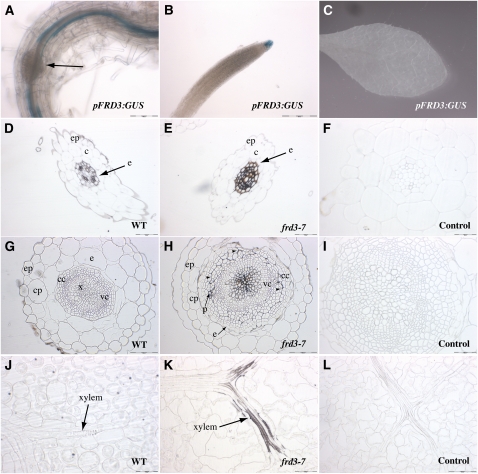
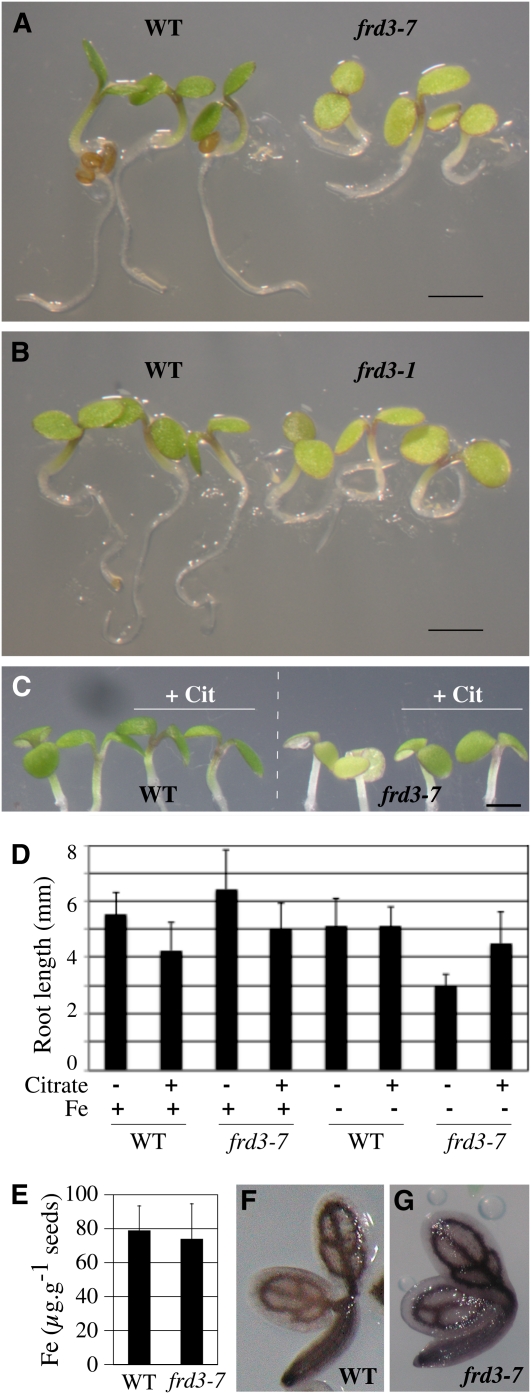
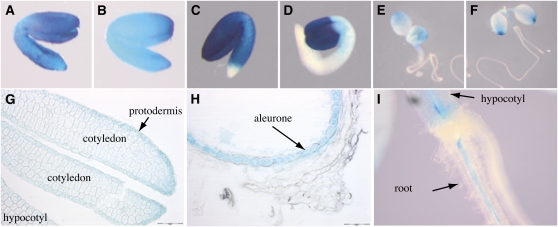
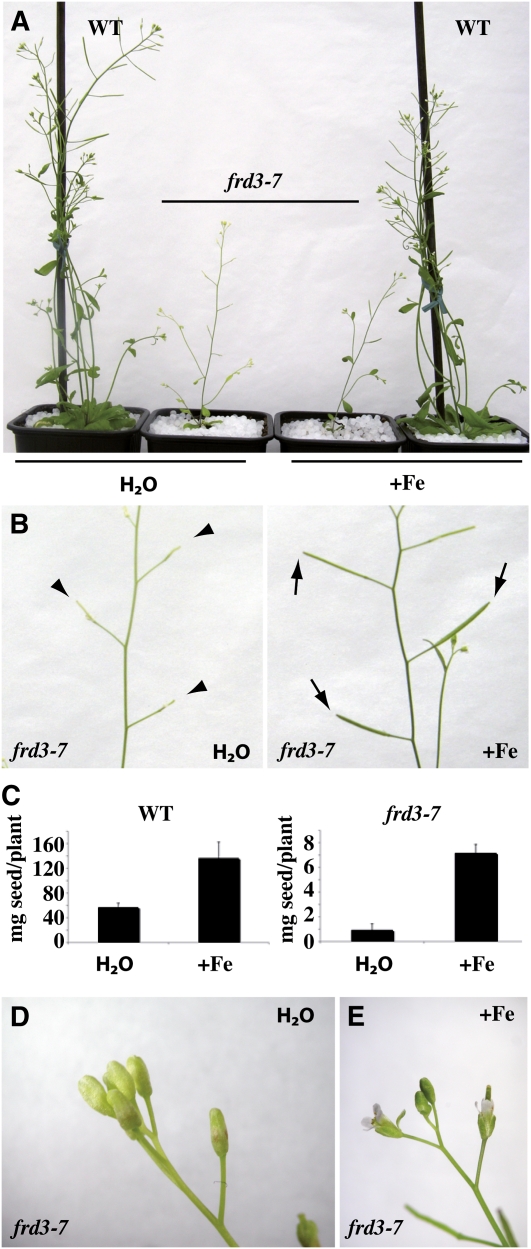
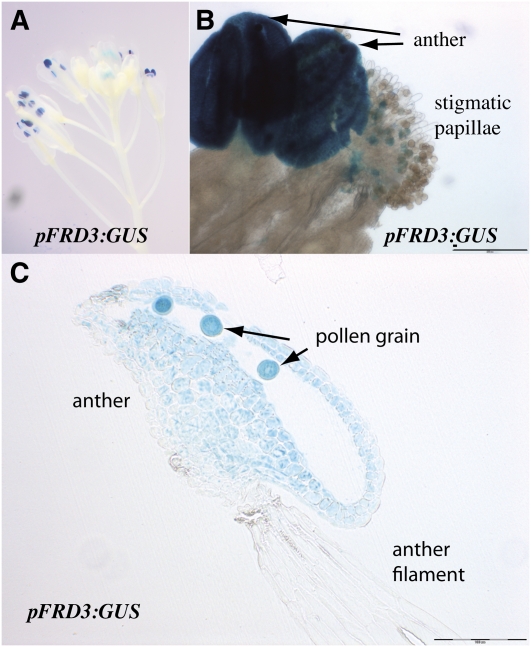
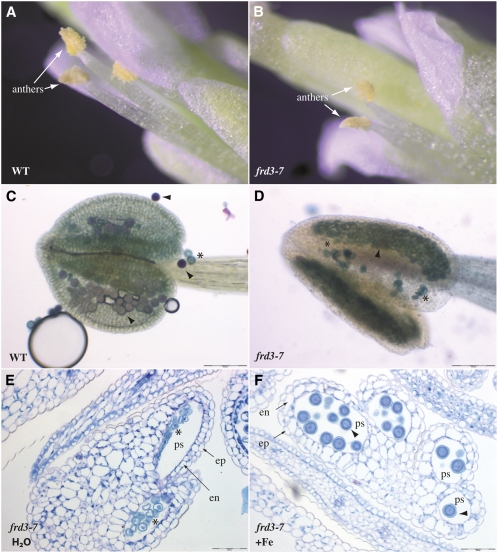
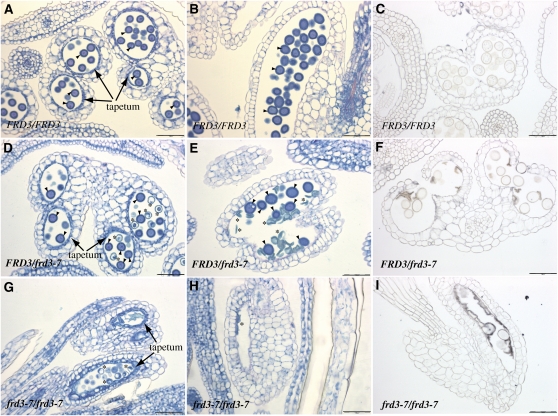
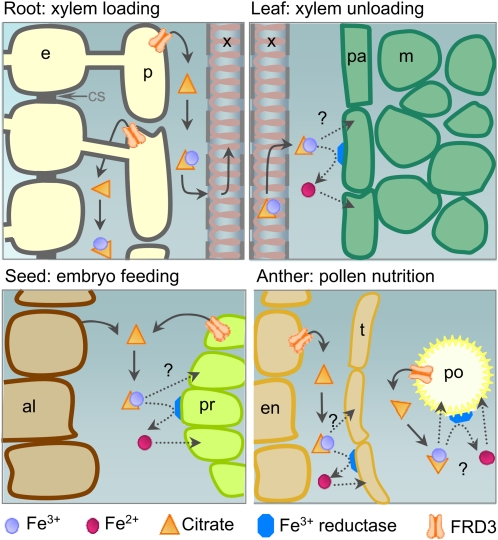
We present data supporting a general role for FERRIC REDICTASE DEFECTIVE3 (FRD3), an efflux transporter of the efficient iron chelator citrate, in maintaining iron homeostasis throughout plant development. In addition to its well-known expression in root, we show that FRD3 is strongly expressed in Arabidopsis thaliana seed and flower. Consistently, frd3 loss-of-function mutants are defective in early germination and are almost completely sterile, both defects being rescued by iron and/or citrate supply. The frd3 fertility defect is caused by pollen abortion and is associated with the male gametophytic expression of FRD3. Iron imaging shows the presence of important deposits of iron on the surface of aborted pollen grains. This points to a role for FRD3 and citrate in proper iron nutrition of embryo and pollen. Based on the findings that iron acquisition in embryo, leaf, and pollen depends on FRD3, we propose that FRD3 mediated-citrate release in the apoplastic space represents an important process by which efficient iron nutrition is achieved between adjacent tissues lacking symplastic connections. These results reveal a physiological role for citrate in the apoplastic transport of iron throughout development, and provide a general model for multicellular organisms in the cell-to-cell transport of iron involving extracellular circulation.
Figures
Similar articles
-
The FRD3-mediated efflux of citrate into the root vasculature is necessary for efficient iron translocation.Plant Physiol. 2007 May;144(1):197-205. doi: 10.1104/pp.107.097162. Epub 2007 Mar 9. Plant Physiol. 2007. PMID: 17351051 Free PMC article.
-
GENERAL CONTROL NONREPRESSED PROTEIN5-Mediated Histone Acetylation of FERRIC REDUCTASE DEFECTIVE3 Contributes to Iron Homeostasis in Arabidopsis.Plant Physiol. 2015 Aug;168(4):1309-20. doi: 10.1104/pp.15.00397. Epub 2015 May 22. Plant Physiol. 2015. PMID: 26002909 Free PMC article.
-
FRD3 controls iron localization in Arabidopsis.Plant Physiol. 2004 Sep;136(1):2523-31. doi: 10.1104/pp.104.045633. Epub 2004 Aug 13. Plant Physiol. 2004. PMID: 15310833 Free PMC article.
-
FRD3, a member of the multidrug and toxin efflux family, controls iron deficiency responses in Arabidopsis.Plant Cell. 2002 Aug;14(8):1787-99. doi: 10.1105/tpc.001495. Plant Cell. 2002. PMID: 12172022 Free PMC article.
-
Intertwined metal homeostasis, oxidative and biotic stress responses in the Arabidopsis frd3 mutant.Plant J. 2020 Apr;102(1):34-52. doi: 10.1111/tpj.14610. Epub 2020 Jan 4. Plant J. 2020. PMID: 31721347
Cited by
-
An RNA-Seq transcriptome analysis of orthophosphate-deficient white lupin reveals novel insights into phosphorus acclimation in plants.Plant Physiol. 2013 Feb;161(2):705-24. doi: 10.1104/pp.112.209254. Epub 2012 Nov 29. Plant Physiol. 2013. PMID: 23197803 Free PMC article.
-
Shoot to root communication is necessary to control the expression of iron-acquisition genes in Strategy I plants.Planta. 2013 Jan;237(1):65-75. doi: 10.1007/s00425-012-1757-0. Epub 2012 Sep 15. Planta. 2013. PMID: 22983673
-
Iron Biofortification in Rice: An Update on Quantitative Trait Loci and Candidate Genes.Front Plant Sci. 2021 May 26;12:647341. doi: 10.3389/fpls.2021.647341. eCollection 2021. Front Plant Sci. 2021. PMID: 34122472 Free PMC article. Review.
-
Nicotianamine Synthase 2 Is Required for Symbiotic Nitrogen Fixation in Medicago truncatula Nodules.Front Plant Sci. 2020 Jan 30;10:1780. doi: 10.3389/fpls.2019.01780. eCollection 2019. Front Plant Sci. 2020. PMID: 32082345 Free PMC article.
-
OPT3 Is a Phloem-Specific Iron Transporter That Is Essential for Systemic Iron Signaling and Redistribution of Iron and Cadmium in Arabidopsis.Plant Cell. 2014 May;26(5):2249-2264. doi: 10.1105/tpc.114.123737. Epub 2014 May 27. Plant Cell. 2014. PMID: 24867923 Free PMC article.
References
-
- Brown J.C. (1966). Fe and Ca uptake as related to root-sap and stem-exudate citrate in soybeans. Physiol. Plant. 19: 968–976
-
- Clough S.J., Bent A.F. (1998). Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16: 735–743 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
