

Expression of the human apoE2 isoform in adipocytes: altered cellular processing and impaired adipocyte lipogenesis
- PMID: 21743035
- PMCID: PMC3151693
- DOI: 10.1194/jlr.M017160
Expression of the human apoE2 isoform in adipocytes: altered cellular processing and impaired adipocyte lipogenesis
Abstract
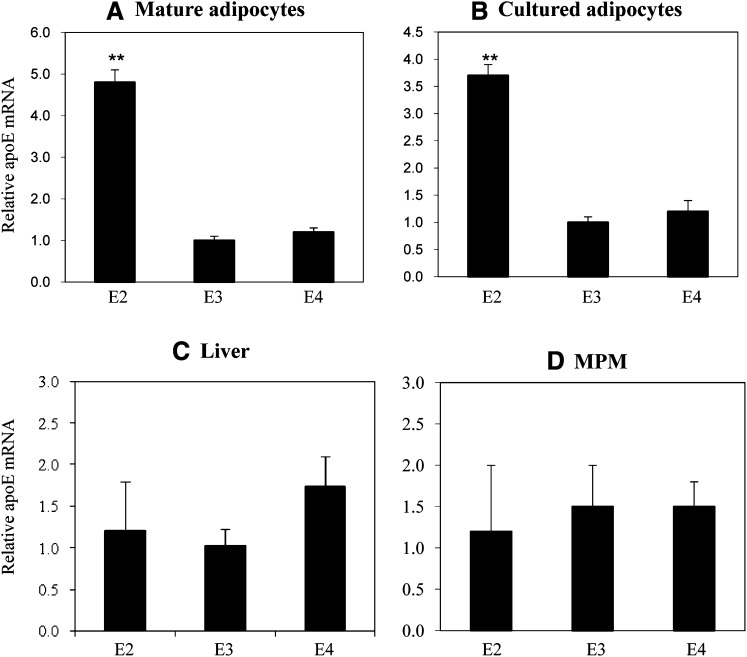
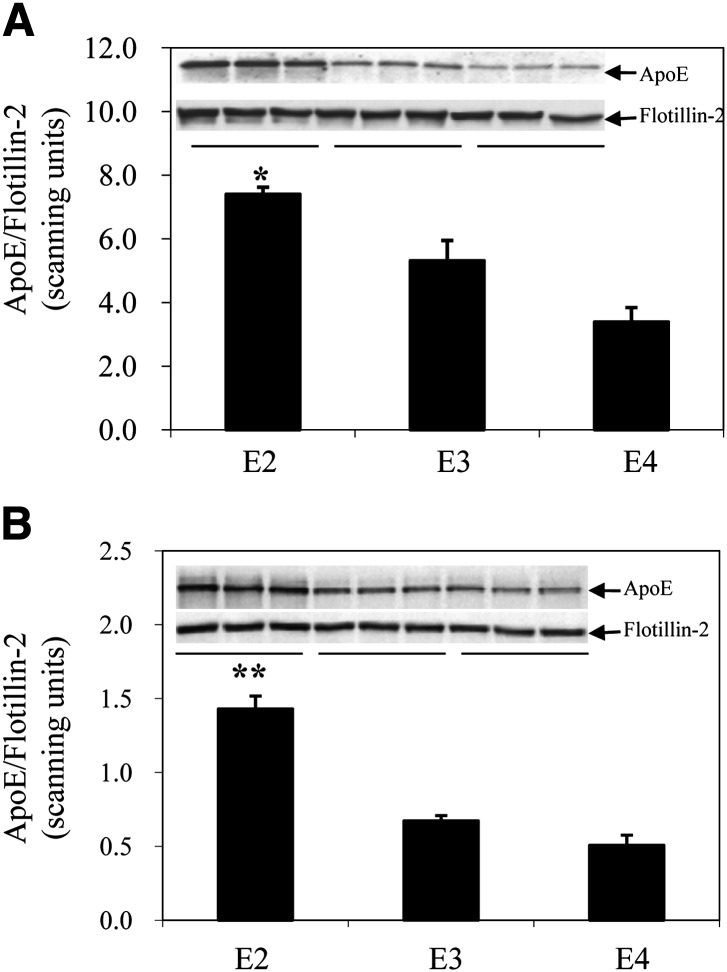
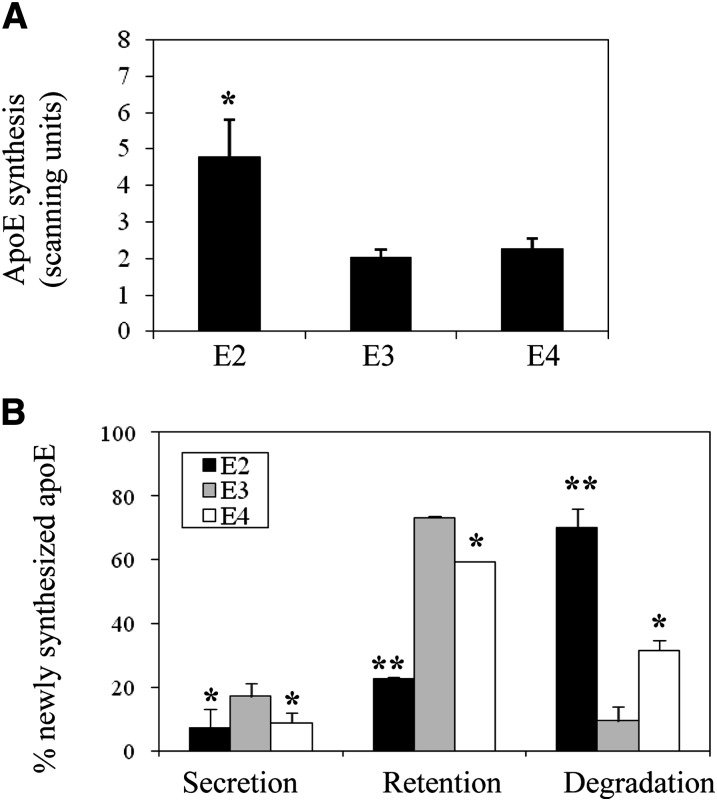
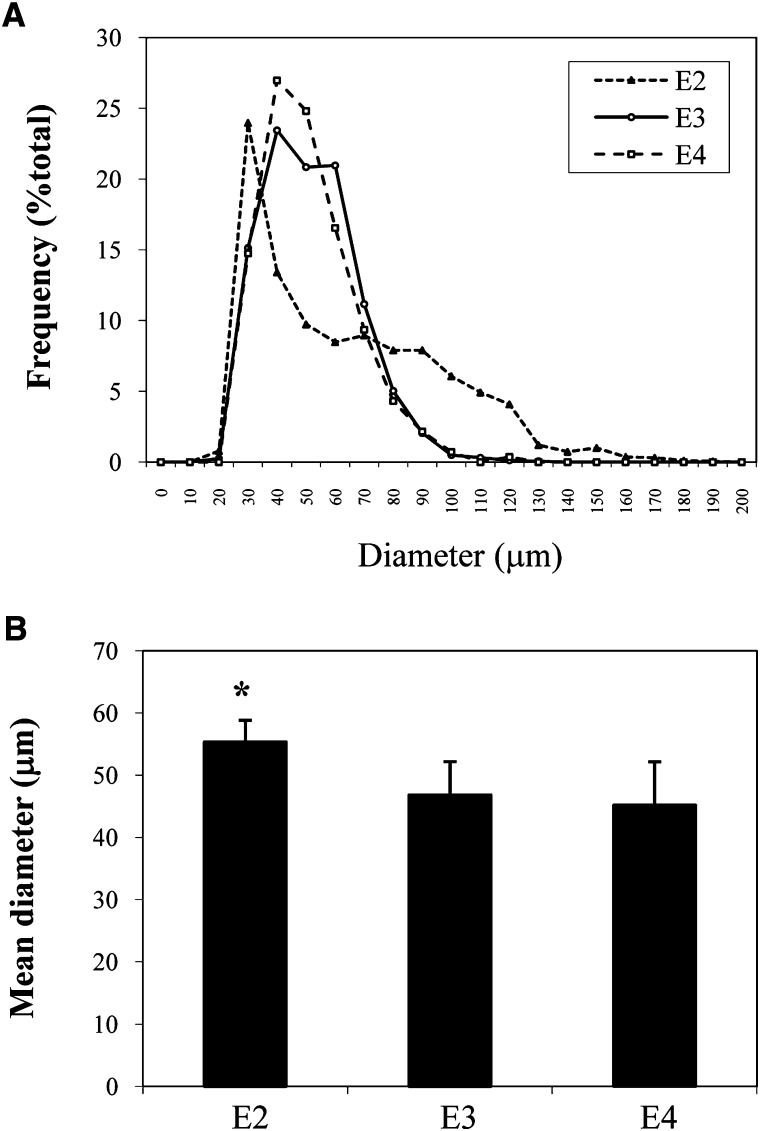
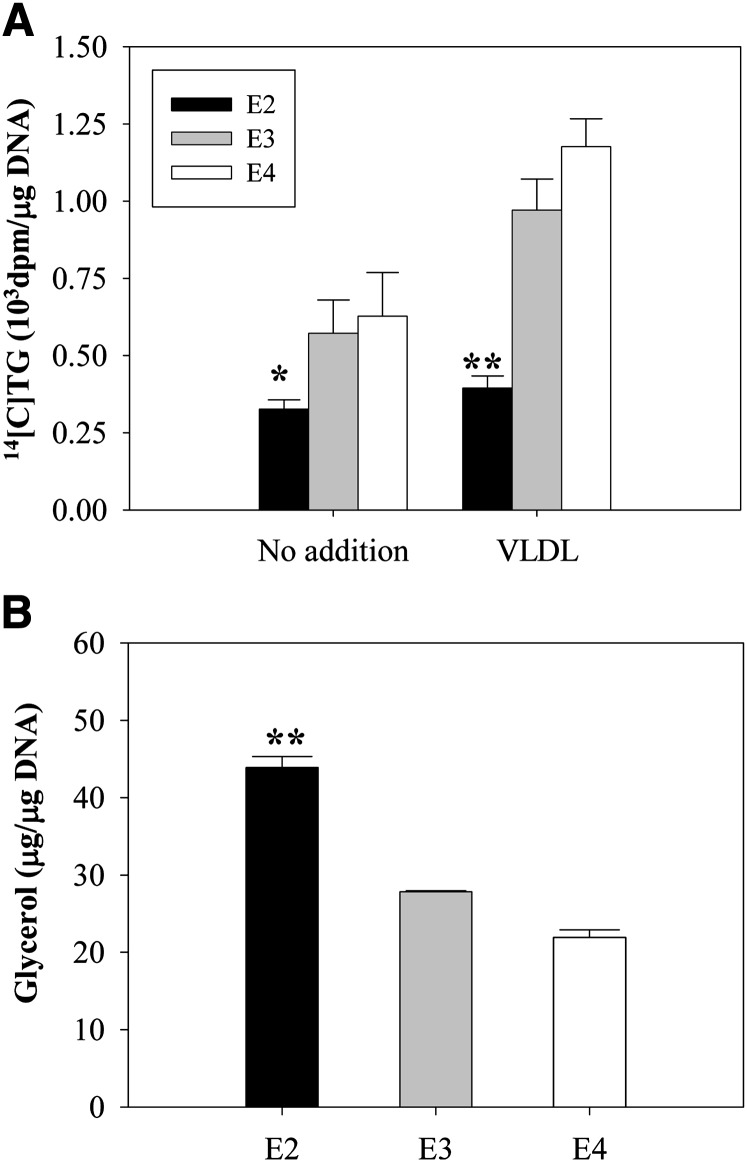
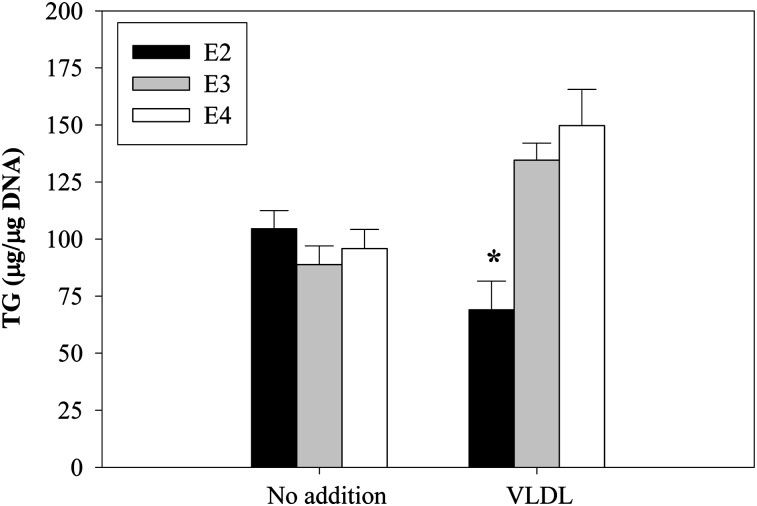
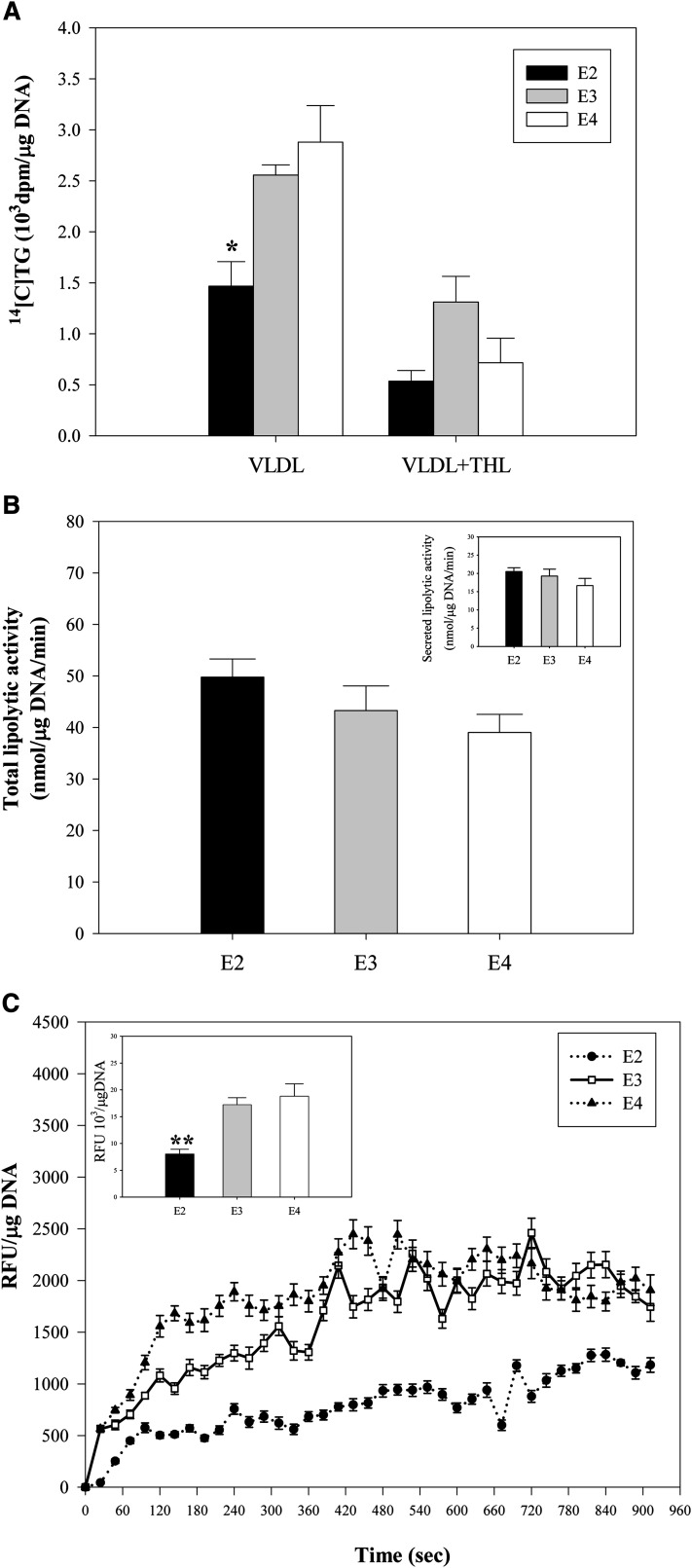
Expression of apoE in adipocytes has been shown to have an important role in modulating adipocyte triglyceride (TG) metabolism and gene expression that is independent of circulating and extracellular apoE. The impact of adipocyte expression of common human apoE isoforms was evaluated using adipocytes harvested from human apoE2, -3, and -4 knock-in mice. Expression of the apoE2 isoform was associated with an increase in adipocyte apoE gene expression and apoE synthesis. Newly synthesized apoE2 was unstable in adipocytes and demonstrated increased degradation and decreased secretion. ApoE2-expressing mice were hyperlipidemic, and had increased size of gonadal fat pads and of adipocytes, compared with apoE3 mice. In isolated cells, however, expression of the apoE2 isoform produced defective lipogenesis and increased TG hydrolysis. Incubation of adipose tissue with apoE3-containing TG-rich lipoproteins resulted in a significant increase in TG in adipose tissue from apoE3 and -E4 mice, but not apoE2 mice. Reduced capacity to internalize FFA as lipogenic substrate contributed to defective lipogenesis. Newly synthesized apoE2 is unstable in adipocytes and results in decreased adipocyte TG synthesis and defective FA uptake. These changes recapitulate those observed in apoE knockout adipocytes and have implications for understanding metabolic disturbances in humans expressing the E2 isoform.
Figures
Similar articles
-
Isoform and tissue dependent impact of apolipoprotein E on adipose tissue metabolic activation: The role of apolipoprotein A1.Biochim Biophys Acta Mol Cell Biol Lipids. 2020 Feb;1865(2):158551. doi: 10.1016/j.bbalip.2019.158551. Epub 2019 Oct 31. Biochim Biophys Acta Mol Cell Biol Lipids. 2020. PMID: 31678510
-
Hyperlipidemia in APOE2 transgenic mice is ameliorated by a truncated apoE variant lacking the C-terminal domain.J Lipid Res. 2003 Feb;44(2):408-14. doi: 10.1194/jlr.M200313-JLR200. Epub 2002 Oct 16. J Lipid Res. 2003. PMID: 12576523
-
Apolipoprotein E2 accentuates postprandial inflammation and diet-induced obesity to promote hyperinsulinemia in mice.Diabetes. 2013 Feb;62(2):382-91. doi: 10.2337/db12-0390. Epub 2012 Sep 6. Diabetes. 2013. PMID: 22961083 Free PMC article.
-
Apolipoprotein E in lipoprotein metabolism, health and cardiovascular disease.Pathology. 2019 Feb;51(2):165-176. doi: 10.1016/j.pathol.2018.11.002. Epub 2018 Dec 28. Pathology. 2019. PMID: 30598326 Review.
-
Apolipoprotein E isoforms and lipoprotein metabolism.IUBMB Life. 2014 Sep;66(9):616-23. doi: 10.1002/iub.1314. IUBMB Life. 2014. PMID: 25328986 Review.
Cited by
-
Features of Lipid Metabolism in Humanized ApoE Knockin Rat Models.Int J Mol Sci. 2021 Jul 31;22(15):8262. doi: 10.3390/ijms22158262. Int J Mol Sci. 2021. PMID: 34361033 Free PMC article.
-
Selective suppression of adipose tissue apoE expression impacts systemic metabolic phenotype and adipose tissue inflammation.J Lipid Res. 2015 Feb;56(2):215-26. doi: 10.1194/jlr.M050567. Epub 2014 Nov 24. J Lipid Res. 2015. PMID: 25421060 Free PMC article.
-
Apolipoprotein E-low density lipoprotein receptor interaction affects spatial memory retention and brain ApoE levels in an isoform-dependent manner.Neurobiol Dis. 2014 Apr;64:150-62. doi: 10.1016/j.nbd.2013.12.016. Epub 2014 Jan 9. Neurobiol Dis. 2014. PMID: 24412220 Free PMC article.
-
Invariant natural killer T cells in adipose tissue: novel regulators of immune-mediated metabolic disease.Cell Mol Life Sci. 2013 Dec;70(24):4711-27. doi: 10.1007/s00018-013-1414-1. Epub 2013 Jul 9. Cell Mol Life Sci. 2013. PMID: 23835837 Free PMC article. Review.
-
Beyond the CNS: The many peripheral roles of APOE.Neurobiol Dis. 2020 May;138:104809. doi: 10.1016/j.nbd.2020.104809. Epub 2020 Feb 19. Neurobiol Dis. 2020. PMID: 32087284 Free PMC article. Review.
References
-
- Ford E. S., Mokdad A. H. 2008. Epidemiology of obesity in the Western Hemisphere. J. Clin. Endocrinol. Metab. 93: S1–S8. - PubMed
-
- Despres J. P., Lemieux I., Bergeron J., Pibarot P., Mathieu P., Larose E., Rodes-Cabau J., Bertrand O. F., Poirier P. 2008. Abdominal obesity and the metabolic syndrome: contribution to global cardiometabolic risk. Arterioscler. Thromb. Vasc. Biol. 28: 1039–1049. - PubMed
-
- Canoy D., Boekholdt S. M., Wareham N., Luben R., Welch A., Bingham S., Buchan I., Day N., Khaw K. T. 2007. Body fat distribution and risk of coronary heart disease in men and women in the European Prospective Investigation Into Cancer and Nutrition in Norfolk cohort: a population-based prospective study. Circulation. 116: 2933–2943. - PubMed
-
- Huang Z. H., Reardon C. A., Mazzone T. 2006. Endogenous apoE expression modulates adipocyte triglyceride content and turnover. Diabetes. 55: 3394–3402. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
