

A chemical probe selectively inhibits G9a and GLP methyltransferase activity in cells
- PMID: 21743462
- PMCID: PMC3184254
- DOI: 10.1038/nchembio.599
A chemical probe selectively inhibits G9a and GLP methyltransferase activity in cells
Erratum in
- Nat Chem Biol. 2011 Sep;7(9):648
- Nat Chem Biol. 2011;7(8):following 574
Abstract
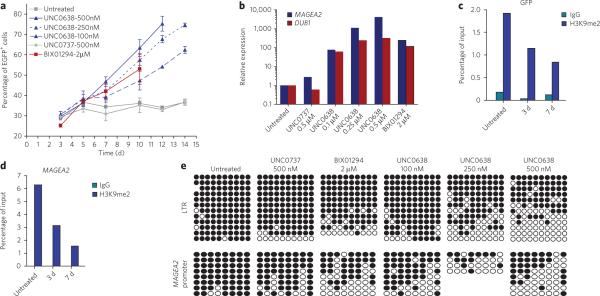
Protein lysine methyltransferases G9a and GLP modulate the transcriptional repression of a variety of genes via dimethylation of Lys9 on histone H3 (H3K9me2) as well as dimethylation of non-histone targets. Here we report the discovery of UNC0638, an inhibitor of G9a and GLP with excellent potency and selectivity over a wide range of epigenetic and non-epigenetic targets. UNC0638 treatment of a variety of cell lines resulted in lower global H3K9me2 levels, equivalent to levels observed for small hairpin RNA knockdown of G9a and GLP with the functional potency of UNC0638 being well separated from its toxicity. UNC0638 markedly reduced the clonogenicity of MCF7 cells, reduced the abundance of H3K9me2 marks at promoters of known G9a-regulated endogenous genes and disproportionately affected several genomic loci encoding microRNAs. In mouse embryonic stem cells, UNC0638 reactivated G9a-silenced genes and a retroviral reporter gene in a concentration-dependent manner without promoting differentiation.
Figures





References
-
- Kouzarides T. Chromatin modifications and their function. Cell. 2007;128:693–705. - PubMed
-
- Martin C, Zhang Y. The diverse functions of histone lysine methylation. Nat. Rev. Mol. Cell Biol. 2005;6:838–849. - PubMed
-
- Jenuwein T, Allis CD. Translating the histone code. Science. 2001;293:1074–1080. - PubMed
-
- Bernstein BE, Meissner A, Lander ES. The mammalian epigenome. Cell. 2007;128:669–681. - PubMed
-
- Gelato KA, Fischle W. Role of histone modifications in defining chromatin structure and function. Biol. Chem. 2008;389:353–363. - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
- R01 MH088413/MH/NIMH NIH HHS/United States
- 199170/CAPMC/ CIHR/Canada
- DP2OD007447/OD/NIH HHS/United States
- R01 MH074127/MH/NIMH NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- R21 HG004535/HG/NHGRI NIH HHS/United States
- DP3DK085698/DK/NIDDK NIH HHS/United States
- IG1-102956/CAPMC/ CIHR/Canada
- DP3 DK085698/DK/NIDDK NIH HHS/United States
- MH074127/MH/NIMH NIH HHS/United States
- HG004535/HG/NHGRI NIH HHS/United States
- 186007/CAPMC/ CIHR/Canada
- DP2 OD007447/OD/NIH HHS/United States
- MH088413/MH/NIMH NIH HHS/United States
- RC1 GM090732/GM/NIGMS NIH HHS/United States
- RC1GM090732/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
