

Protein targeting and degradation are coupled for elimination of mislocalized proteins
- PMID: 21743475
- PMCID: PMC3150218
- DOI: 10.1038/nature10181
Protein targeting and degradation are coupled for elimination of mislocalized proteins
Abstract
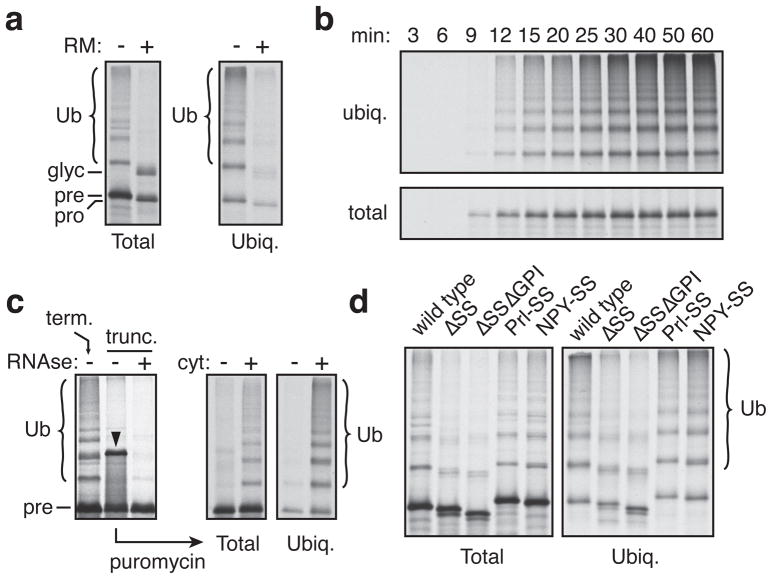
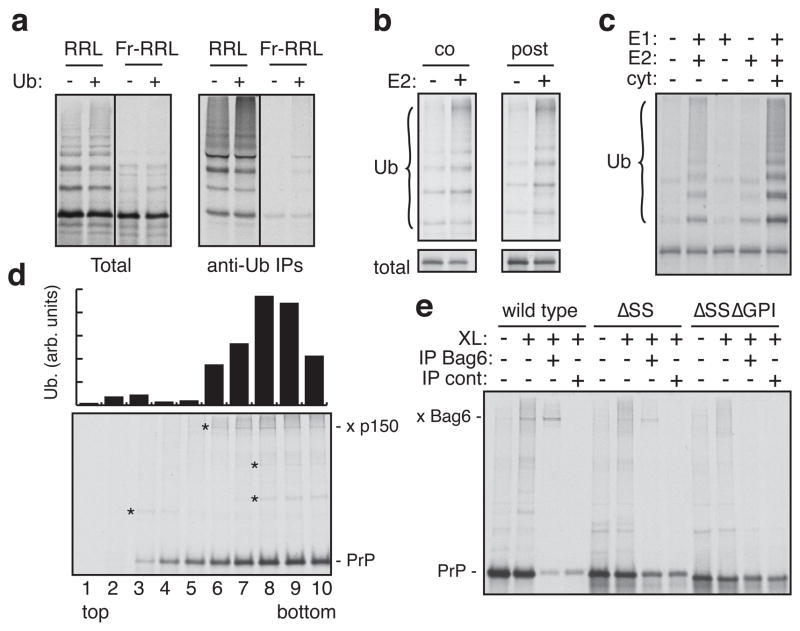
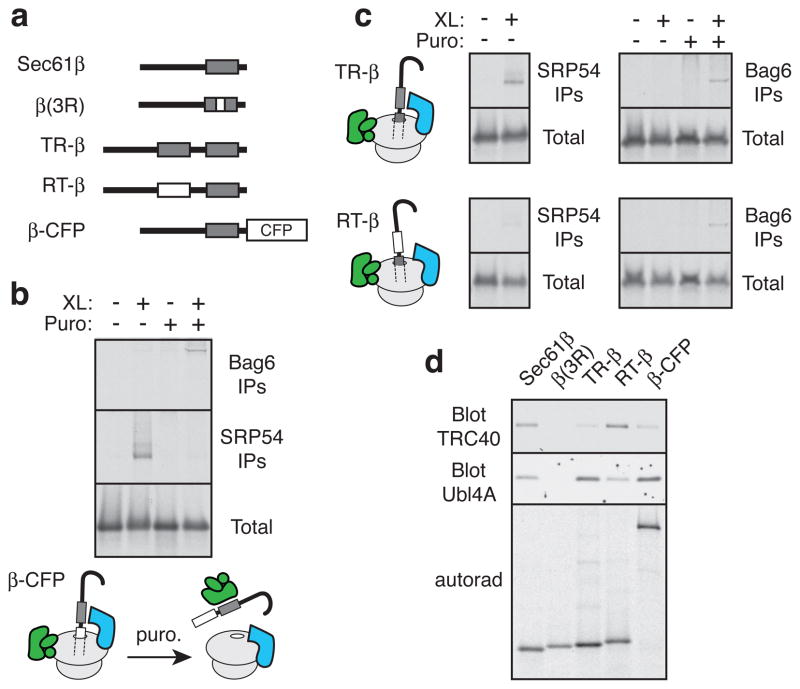
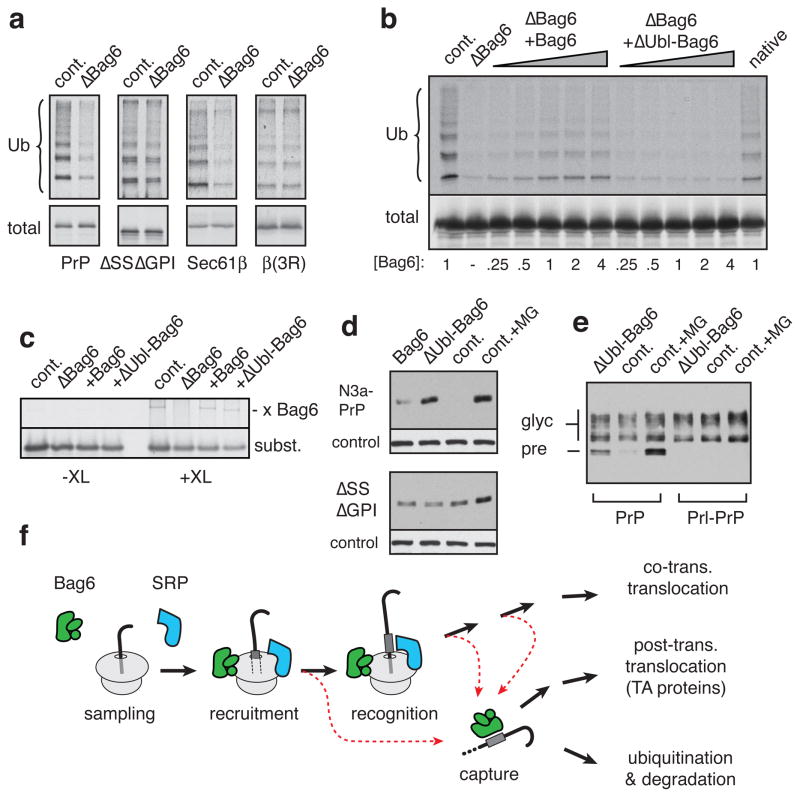
A substantial proportion of the genome encodes membrane proteins that are delivered to the endoplasmic reticulum by dedicated targeting pathways. Membrane proteins that fail targeting must be rapidly degraded to avoid aggregation and disruption of cytosolic protein homeostasis. The mechanisms of mislocalized protein (MLP) degradation are unknown. Here we reconstitute MLP degradation in vitro to identify factors involved in this pathway. We find that nascent membrane proteins tethered to ribosomes are not substrates for ubiquitination unless they are released into the cytosol. Their inappropriate release results in capture by the Bag6 complex, a recently identified ribosome-associating chaperone. Bag6-complex-mediated capture depends on the presence of unprocessed or non-inserted hydrophobic domains that distinguish MLPs from potential cytosolic proteins. A subset of these Bag6 complex 'clients' are transferred to TRC40 for insertion into the membrane, whereas the remainder are rapidly ubiquitinated. Depletion of the Bag6 complex selectively impairs the efficient ubiquitination of MLPs. Thus, by its presence on ribosomes that are synthesizing nascent membrane proteins, the Bag6 complex links targeting and ubiquitination pathways. We propose that such coupling allows the fast tracking of MLPs for degradation without futile engagement of the cytosolic folding machinery.
Figures
Comment in
-
BAG6 'mops up' mislocalized proteins.Nat Rev Mol Cell Biol. 2011 Jul 27;12(9):550. doi: 10.1038/nrm3167. Nat Rev Mol Cell Biol. 2011. PMID: 21792219 No abstract available.
References
-
- Cross BC, Sinning I, Luirink J, High S. Delivering proteins for export from the cytosol. Nat Rev Mol Cell Biol. 2009;10:255–264. - PubMed
-
- Kim SJ, Mitra D, Salerno JR, Hegde RS. Signal sequences control gating of the protein translocation channel in a substrate-specific manner. Dev Cell. 2002;2:207–217. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
