

Ank3-dependent SVZ niche assembly is required for the continued production of new neurons
- PMID: 21745638
- PMCID: PMC3134799
- DOI: 10.1016/j.neuron.2011.05.029
Ank3-dependent SVZ niche assembly is required for the continued production of new neurons
Abstract
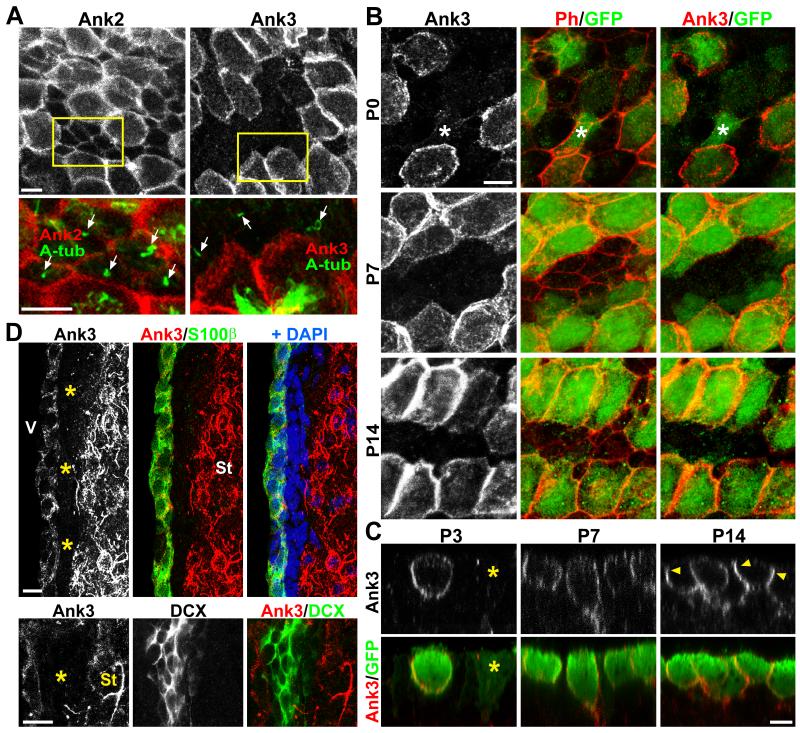
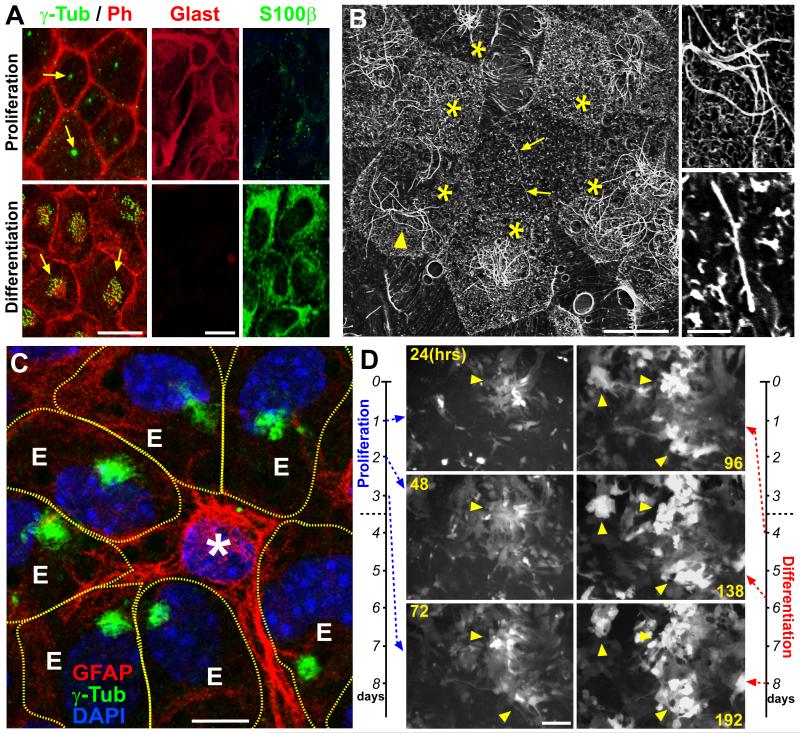
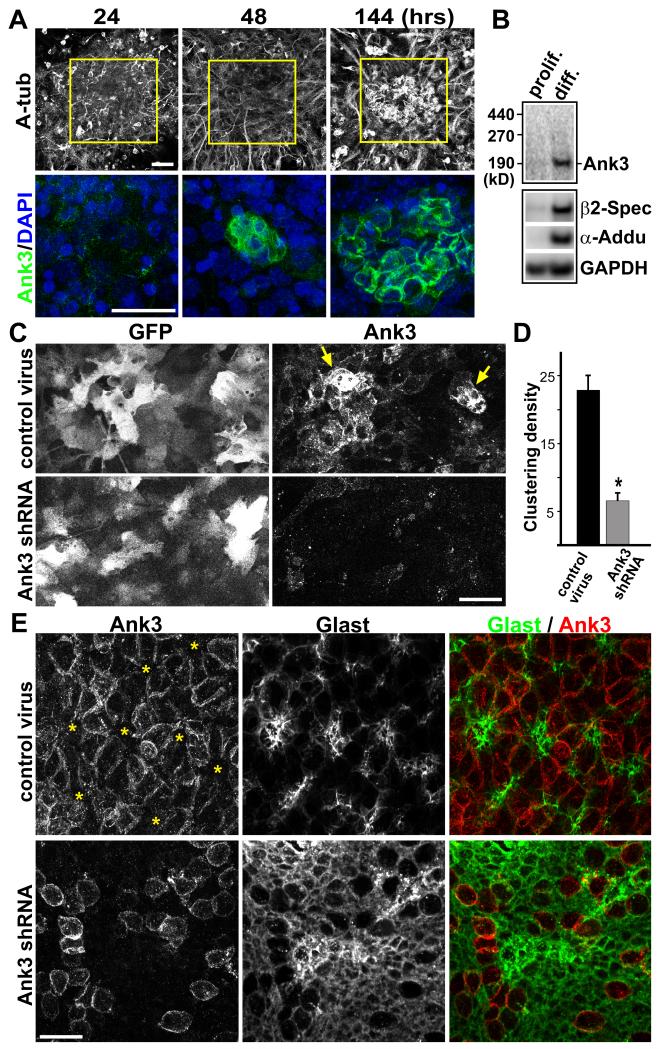
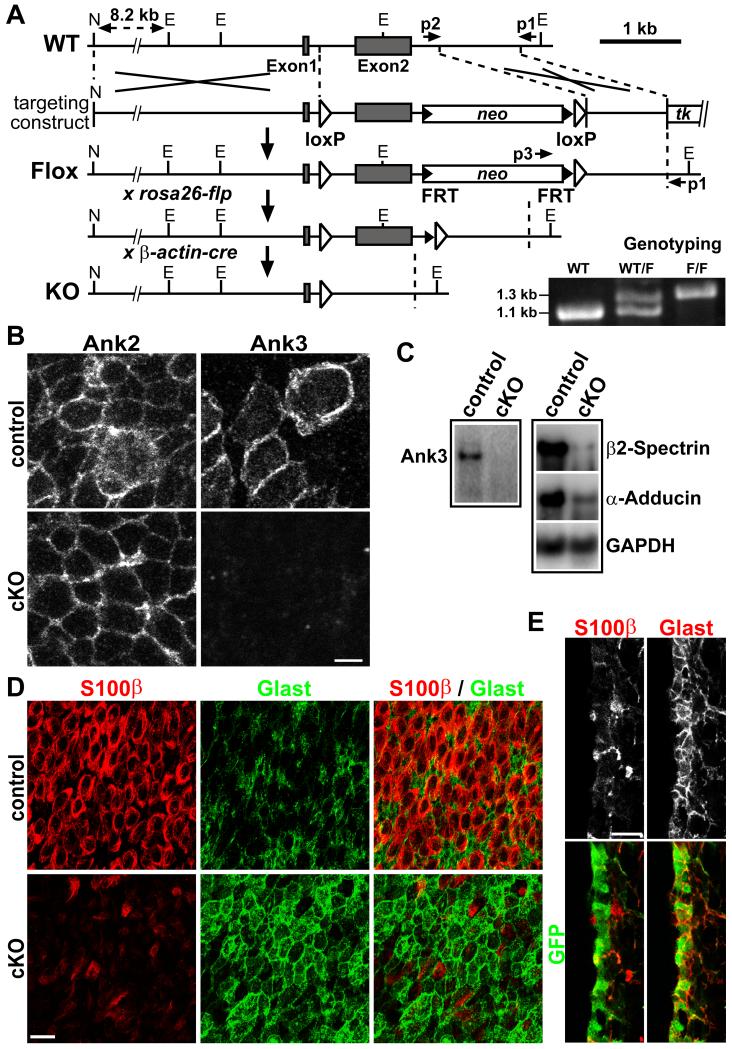
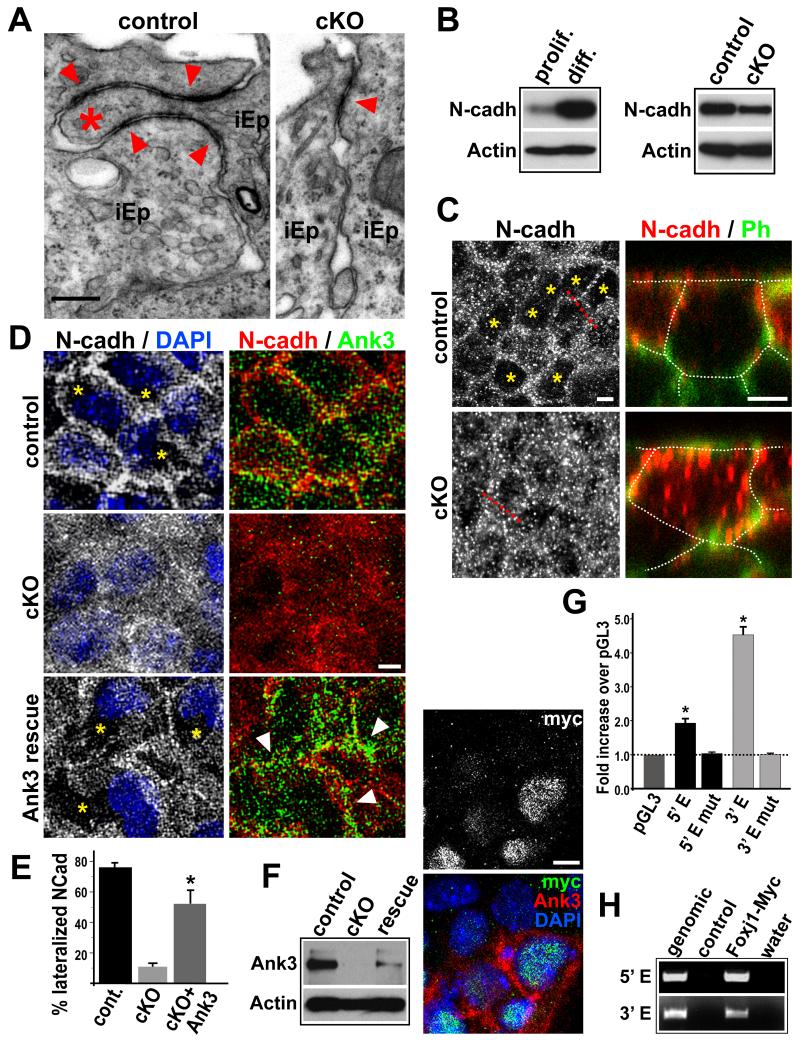
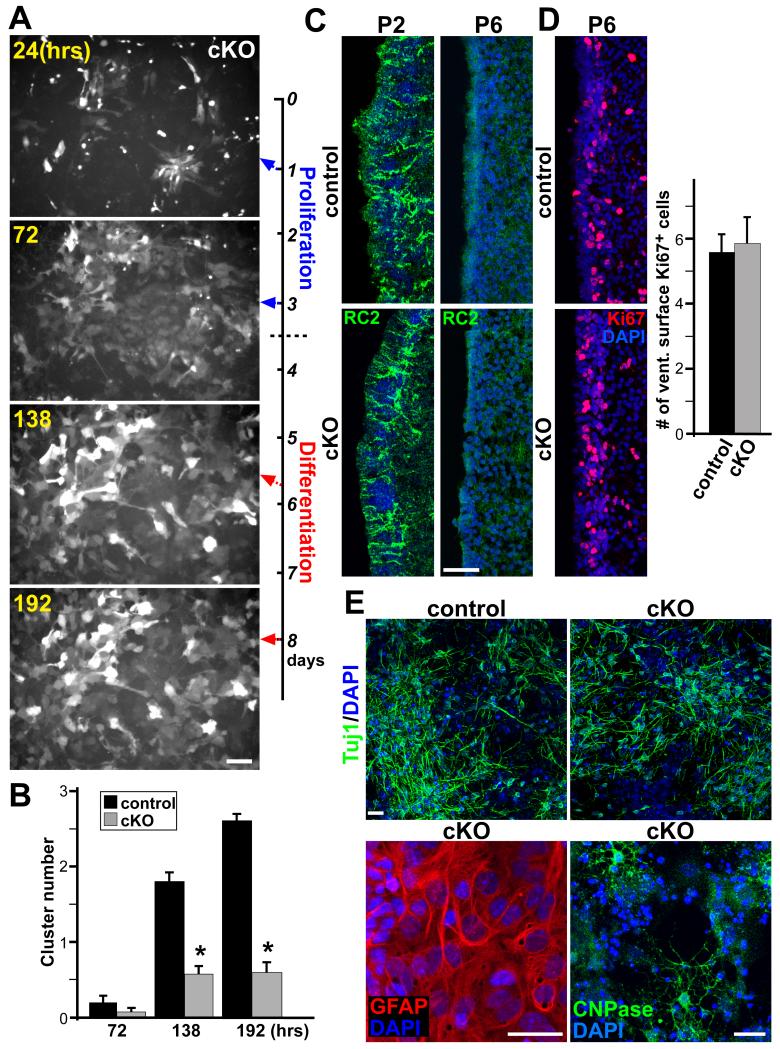
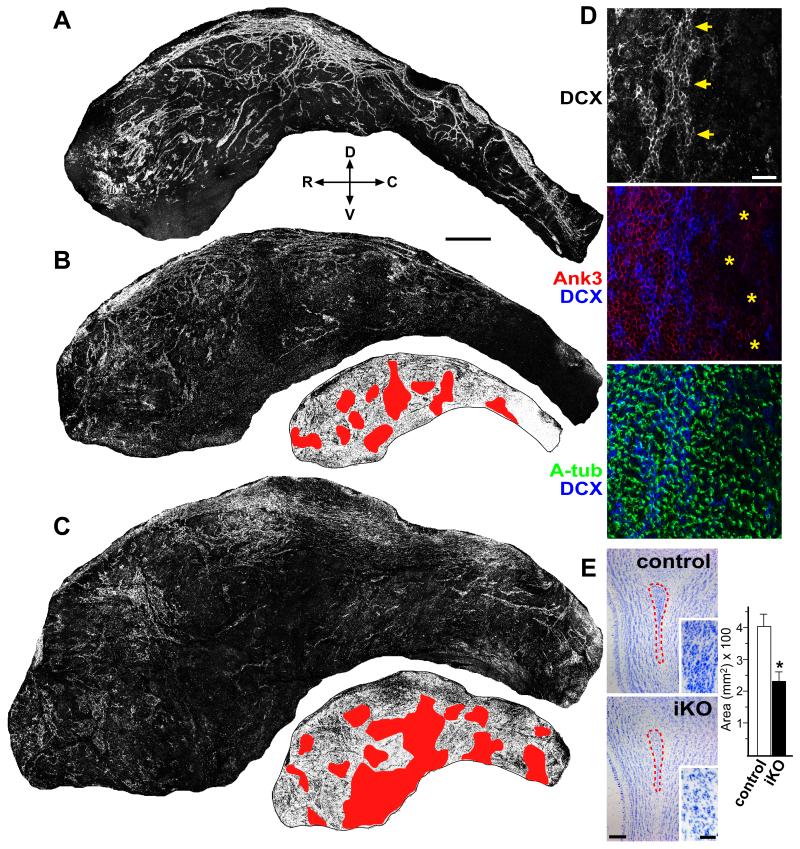
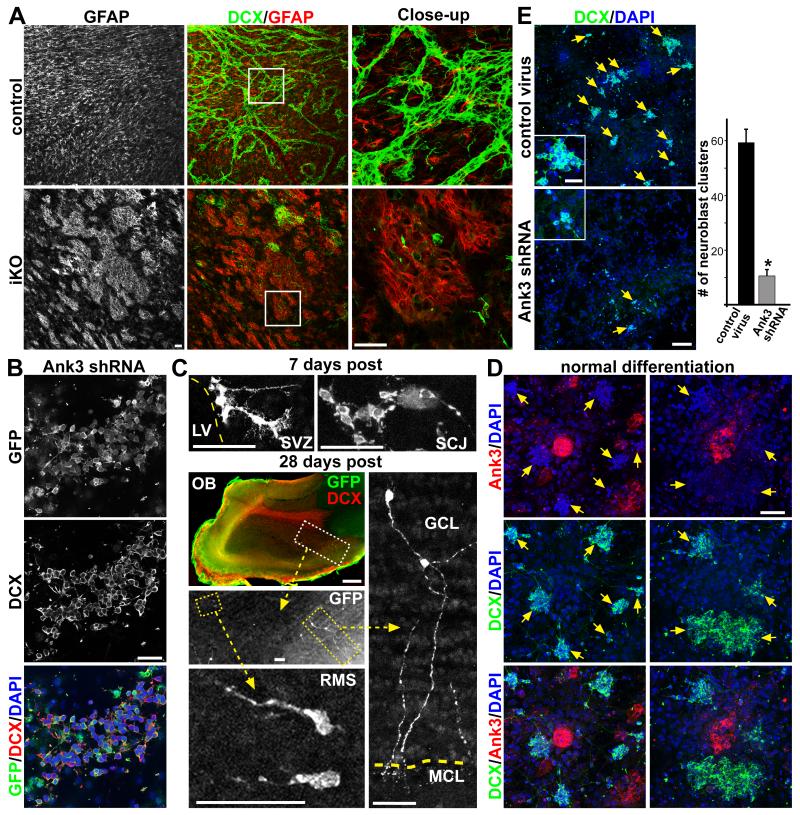
The rodent subventricular/subependymal zone (SVZ/SEZ) houses neural stem cells (NSCs) that generate olfactory bulb interneurons. It is unclear how the SVZ environment sustains neuronal production into adulthood. We discovered that the adapter molecule Ankyrin-3 (Ank3) is specifically upregulated in ventricular progenitors destined to become ependymal cells, but not in NSCs, and is required for SVZ niche assembly through progenitor lateral adhesion. Furthermore, we found that Ank3 expression is controlled by Foxj1, a transcriptional regulator of multicilia formation, and genetic deletion of this pathway led to complete loss of SVZ niche structure. Interestingly, radial glia continued to transition into postnatal NSCs without this niche. However, inducible deletion of Foxj1-Ank3 from mature SVZ ependyma resulted in dramatic depletion of neurogenesis. Targeting a pathway regulating ependymal organization/assembly and showing its requirement for new neuron production, our results have important implications for environmental control of adult neurogenesis and harvesting NSCs for replacement therapy.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Alvarez-Buylla A, Lim DA. For the long run: maintaining germinal niches in the adult brain. Neuron. 2004;41:683–686. - PubMed
-
- Bejerano G, Siepel AC, Kent WJ, Haussler D. Computational screening of conserved genomic DNA in search of functional noncoding elements. Nat Methods. 2005;2:535–545. - PubMed
-
- Bennett V, Healy J. Organizing the fluid membrane bilayer: diseases linked to spectrin and ankyrin. Trends Mol Med. 2008;14:28–36. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
