

Relation of addiction genes to hypothalamic gene changes subserving genesis and gratification of a classic instinct, sodium appetite
- PMID: 21746918
- PMCID: PMC3145743
- DOI: 10.1073/pnas.1109199108
Relation of addiction genes to hypothalamic gene changes subserving genesis and gratification of a classic instinct, sodium appetite
Abstract
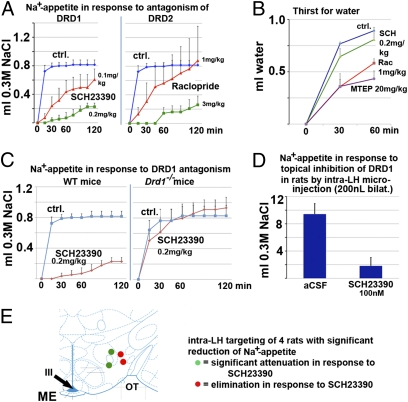
Sodium appetite is an instinct that involves avid specific intention. It is elicited by sodium deficiency, stress-evoked adrenocorticotropic hormone (ACTH), and reproduction. Genome-wide microarrays in sodium-deficient mice or after ACTH infusion showed up-regulation of hypothalamic genes, including dopamine- and cAMP-regulated neuronal phosphoprotein 32 kDa (DARPP-32), dopamine receptors-1 and -2, α-2C- adrenoceptor, and striatally enriched protein tyrosine phosphatase (STEP). Both DARPP-32 and neural plasticity regulator activity-regulated cytoskeleton associated protein (ARC) were up-regulated in lateral hypothalamic orexinergic neurons by sodium deficiency. Administration of dopamine D1 (SCH23390) and D2 receptor (raclopride) antagonists reduced gratification of sodium appetite triggered by sodium deficiency. SCH23390 was specific, having no effect on osmotic-induced water drinking, whereas raclopride also reduced water intake. D1 receptor KO mice had normal sodium appetite, indicating compensatory regulation. Appetite was insensitive to SCH23390, confirming the absence of off-target effects. Bilateral microinjection of SCH23390 (100 nM in 200 nL) into rats' lateral hypothalamus greatly reduced sodium appetite. Gene set enrichment analysis in hypothalami of mice with sodium appetite showed significant enrichment of gene sets previously linked to addiction (opiates and cocaine). This finding of concerted gene regulation was attenuated on gratification with perplexingly rapid kinetics of only 10 min, anteceding significant absorption of salt from the gut. Salt appetite and hedonic liking of salt taste have evolved over >100 million y (e.g., being present in Metatheria). Drugs causing pleasure and addiction are comparatively recent and likely reflect usurping of evolutionary ancient systems with high survival value by the gratification of contemporary hedonic indulgences. Our findings outline a molecular logic for instinctive behavior encoded by the brain with possible important translational-medical implications.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Comment in
-
Hypothalamic gene changes in response to salt appetite.Kidney Int. 2011 Oct;80(8):791. doi: 10.1038/ki.2011.325. Kidney Int. 2011. PMID: 21960166 No abstract available.
References
-
- Denton D. The Hunger for Salt. An Anthropological, Physiological and Medical Analysis. Berlin: Springer; 1983.
-
- Adolph EF, Dill DB. Observations on water metabolism in the desert. Am J Physiol. 1938;123:369–378.
-
- Schmidt-Nielsen K, Schmidt-Nielsen B, Jarnum SA, Houpt TR. Body temperature of the camel and its relation to water economy. Am J Physiol. 1957;188:103–112. - PubMed
-
- Denton D, McBurnie M, Ong F, Osborne P, Tarjan E. Na deficiency and other physiological influences on voluntary Na intake of BALB/c mice. Am J Physiol. 1988;255:R1025–R1034. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
