

BDNF facilitates L-LTP maintenance in the absence of protein synthesis through PKMζ
- PMID: 21747912
- PMCID: PMC3126837
- DOI: 10.1371/journal.pone.0021568
BDNF facilitates L-LTP maintenance in the absence of protein synthesis through PKMζ
Abstract
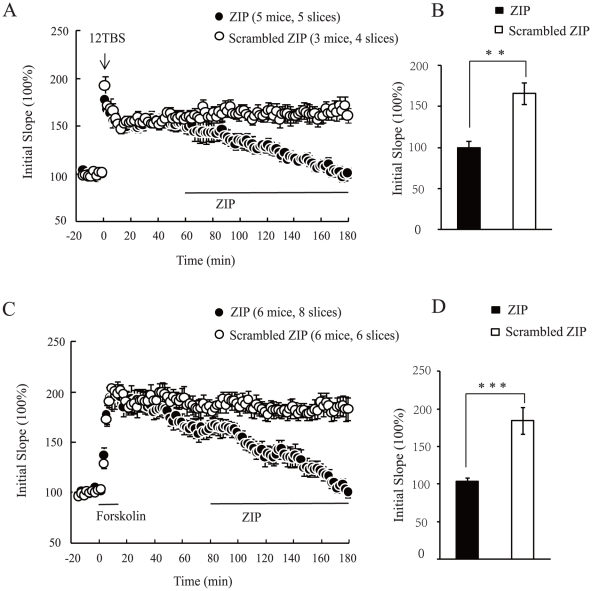
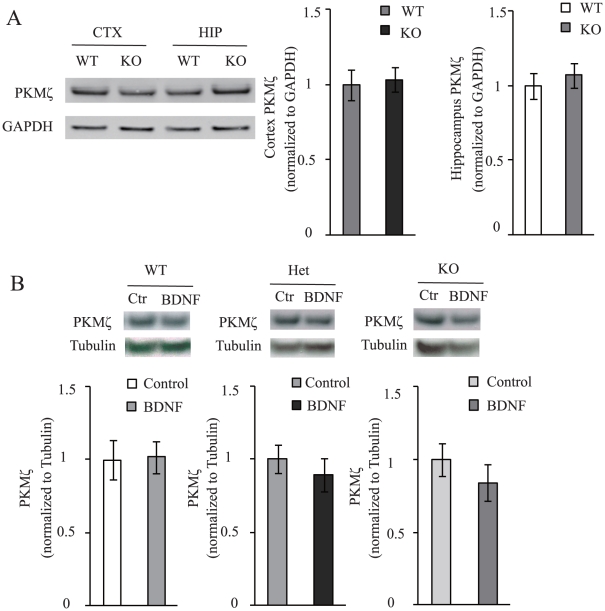
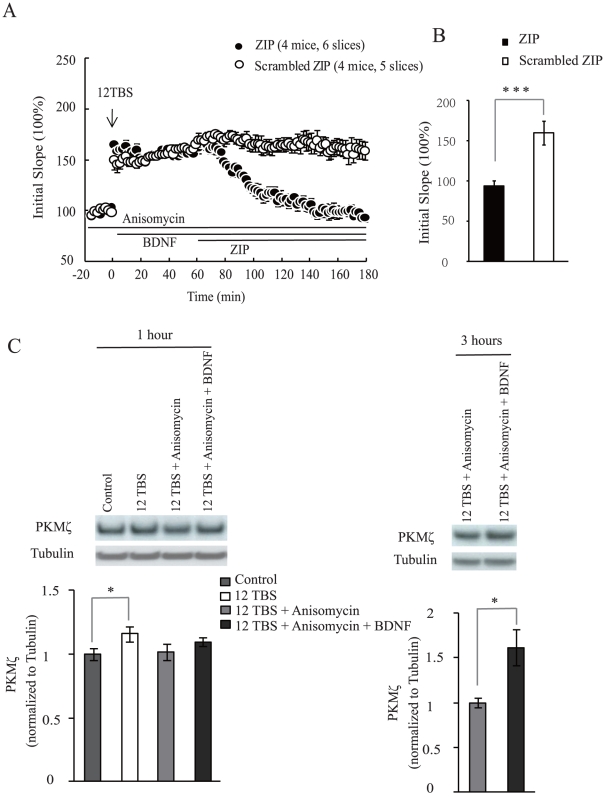
Late-phase long term potentiation (L-LTP) is thought to be the cellular basis for long-term memory (LTM). While LTM as well as L-LTP is known to depend on transcription and translation, it is unclear why brain-derived neurotrophic factor (BDNF) could sustain L-LTP when protein synthesis is inhibited. The persistently active protein kinase ζ (PKMζ) is the only molecule implicated in perpetuating L-LTP maintenance. Here, in mouse acute brain slices, we show that inhibition of PKMζ reversed BDNF-dependent form of L-LTP. While BDNF did not alter the steady-state level of PKMζ, BDNF together with the L-LTP inducing theta-burst stimulation (TBS) increased PKMζ level even without protein synthesis. Finally, in the absence of de novo protein synthesis, BDNF maintained TBS-induced PKMζ at a sufficient level. These results suggest that BDNF sustains L-LTP through PKMζ in a protein synthesis-independent manner, revealing an unexpected link between BDNF and PKMζ.
Conflict of interest statement
Figures
Similar articles
-
PKMzeta, LTP maintenance, and the dynamic molecular biology of memory storage.Prog Brain Res. 2008;169:27-40. doi: 10.1016/S0079-6123(07)00002-7. Prog Brain Res. 2008. PMID: 18394466 Review.
-
BDNF mechanisms in late LTP formation: A synthesis and breakdown.Neuropharmacology. 2014 Jan;76 Pt C:664-76. doi: 10.1016/j.neuropharm.2013.06.024. Epub 2013 Jul 2. Neuropharmacology. 2014. PMID: 23831365 Review.
-
Overexpression of Protein Kinase Mζ in the Hippocampus Enhances Long-Term Potentiation and Long-Term Contextual But Not Cued Fear Memory in Rats.J Neurosci. 2016 Apr 13;36(15):4313-24. doi: 10.1523/JNEUROSCI.3600-15.2016. J Neurosci. 2016. PMID: 27076427 Free PMC article.
-
Compensation for PKMζ in long-term potentiation and spatial long-term memory in mutant mice.Elife. 2016 May 17;5:e14846. doi: 10.7554/eLife.14846. Elife. 2016. PMID: 27187150 Free PMC article.
-
Persistent increases of PKMζ in memory-activated neurons trace LTP maintenance during spatial long-term memory storage.Eur J Neurosci. 2021 Oct;54(8):6795-6814. doi: 10.1111/ejn.15137. Epub 2021 Mar 28. Eur J Neurosci. 2021. PMID: 33540466 Free PMC article.
Cited by
-
Phosphodiesterase 4 inhibition after retrieval switches the memory fate favoring extinction instead of reconsolidation.Sci Rep. 2023 Nov 21;13(1):20384. doi: 10.1038/s41598-023-47717-1. Sci Rep. 2023. PMID: 37990053 Free PMC article.
-
Is plasticity of synapses the mechanism of long-term memory storage?NPJ Sci Learn. 2019 Jul 2;4:9. doi: 10.1038/s41539-019-0048-y. eCollection 2019. NPJ Sci Learn. 2019. PMID: 31285847 Free PMC article. Review.
-
Molecular constraints on synaptic tagging and maintenance of long-term potentiation: a predictive model.PLoS Comput Biol. 2012;8(8):e1002620. doi: 10.1371/journal.pcbi.1002620. Epub 2012 Aug 2. PLoS Comput Biol. 2012. PMID: 22876169 Free PMC article.
-
Long-lasting LTP requires neither repeated trains for its induction nor protein synthesis for its development.PLoS One. 2012;7(7):e40823. doi: 10.1371/journal.pone.0040823. Epub 2012 Jul 11. PLoS One. 2012. PMID: 22792408 Free PMC article.
-
What does LTP tell us about the roles of CaMKII and PKMζ in memory?Mol Brain. 2018 Dec 28;11(1):77. doi: 10.1186/s13041-018-0420-5. Mol Brain. 2018. PMID: 30593289 Free PMC article. Review.
References
-
- Martin SJ, Grimwood PD, Morris RG. Synaptic plasticity and memory: an evaluation of the hypothesis. Annu Rev Neurosci. 2000;23:649–711. - PubMed
-
- Neves G, Cooke SF, Bliss TV. Synaptic plasticity, memory and the hippocampus: a neural network approach to causality. Nat Rev Neurosci. 2008;9:65–75. - PubMed
-
- Bliss TV, Collingridge GL. A synaptic model of memory: long-term potentiation in the hippocampus. Nature. 1993;361:31–39. - PubMed
-
- Davis HP, Squire LR. Protein synthesis and memory: a review. Psychol Bull. 1984;96:518–559. - PubMed
-
- Pang PT, Teng HK, Zaitsev E, Woo NT, Sakata K, et al. Cleavage of proBDNF by tPA/plasmin is essential for long-term hippocampal plasticity. Science. 2004;306:487–491. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
