

Metabolic programming during lactation stimulates renal Na+ transport in the adult offspring due to an early impact on local angiotensin II pathways
- PMID: 21747933
- PMCID: PMC3128598
- DOI: 10.1371/journal.pone.0021232
Metabolic programming during lactation stimulates renal Na+ transport in the adult offspring due to an early impact on local angiotensin II pathways
Abstract
Background: Several studies have correlated perinatal malnutrition with diseases in adulthood, giving support to the programming hypothesis. In this study, the effects of maternal undernutrition during lactation on renal Na(+)-transporters and on the local angiotensin II (Ang II) signaling cascade in rats were investigated.
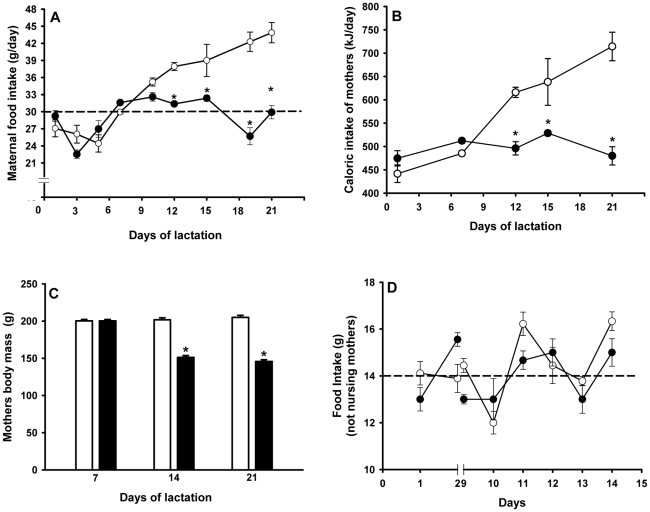
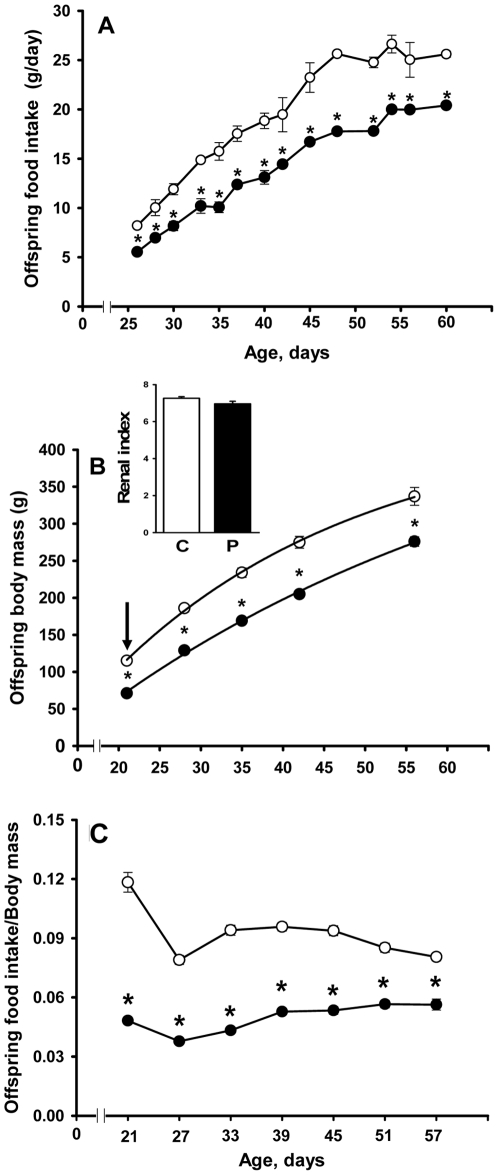
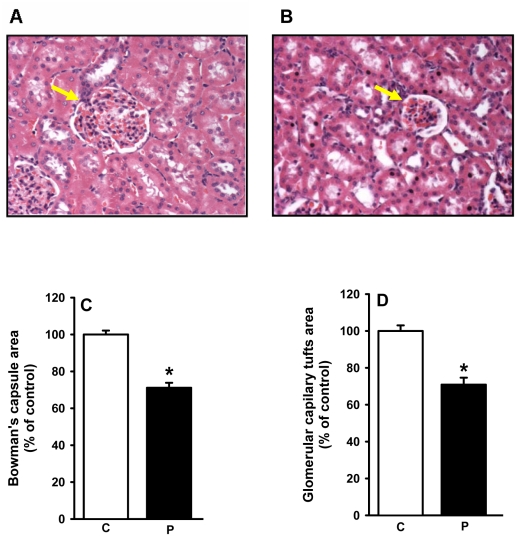
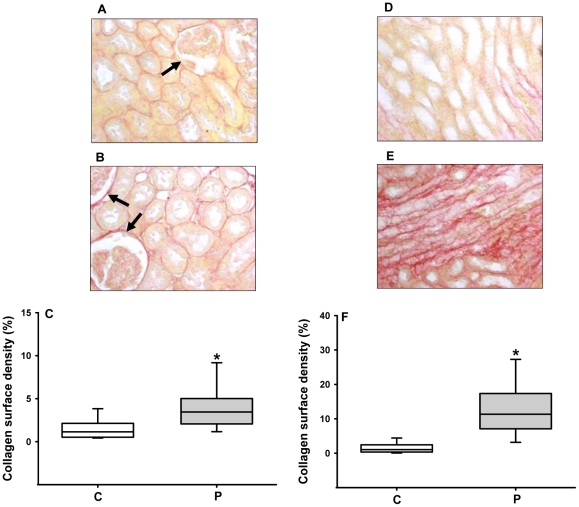
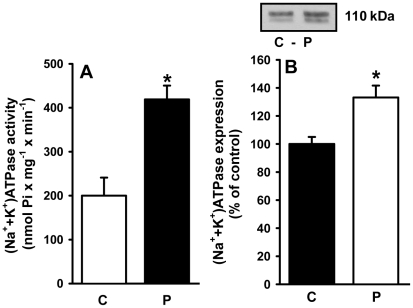
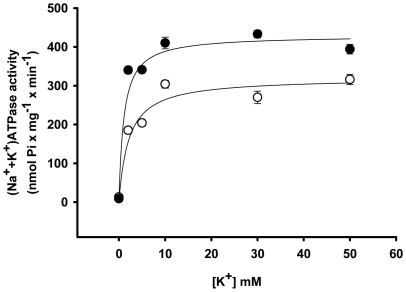
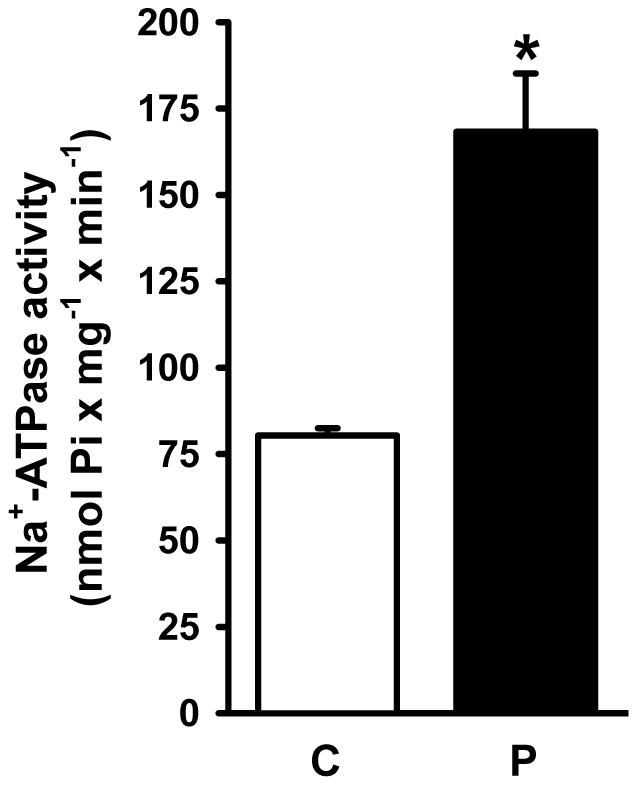
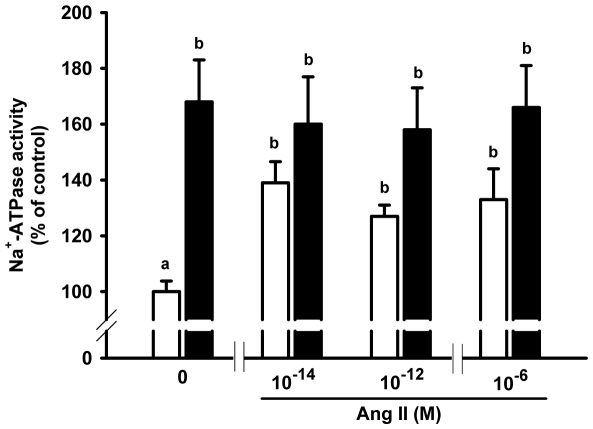
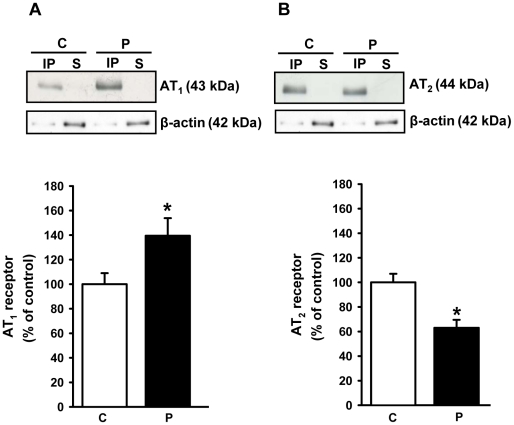
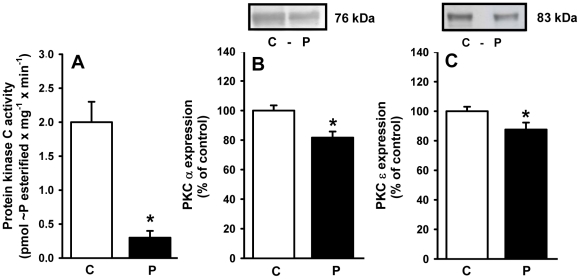
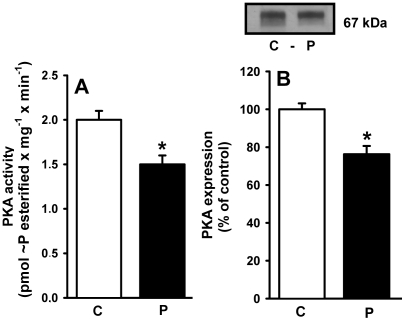
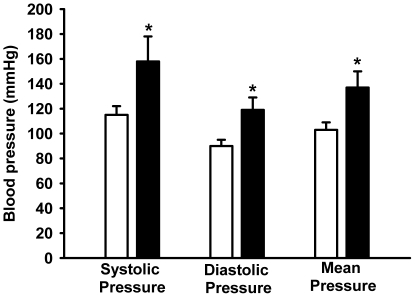
Methodology/principal findings: Female rats received a hypoproteic diet (8% protein) throughout lactation. Control and programmed offspring consumed a diet containing 20% protein after weaning. Programming caused a decrease in the number of nephrons (35%), in the area of the Bowman's capsule (30%) and the capillary tuft (30%), and increased collagen deposition in the cortex and medulla (by 175% and 700%, respectively). In programmed rats the expression of (Na(+)+K(+))ATPase in proximal tubules increased by 40%, but its activity was doubled owing to a threefold increase in affinity for K(+). Programming doubled the ouabain-insensitive Na(+)-ATPase activity with loss of its physiological response to Ang II, increased the expression of AT(1) and decreased the expression of AT(2) receptors), and caused a pronounced inhibition (90%) of protein kinase C activity with decrease in the expression of the α (24%) and ε (13%) isoforms. Activity and expression of cyclic AMP-dependent protein kinase decreased in the same proportion as the AT(2) receptors (30%). In vivo studies at 60 days revealed an increased glomerular filtration rate (GFR) (70%), increased Na(+) excretion (80%) and intense proteinuria (increase of 400% in protein excretion). Programmed rats, which had normal arterial pressure at 60 days, became hypertensive by 150 days.
Conclusions/significance: Maternal protein restriction during lactation results in alterations in GFR, renal Na(+) handling and in components of the Ang II-linked regulatory pathway of renal Na(+) reabsorption. At the molecular level, they provide a framework for understanding how metabolic programming of renal mechanisms contributes to the onset of hypertension in adulthood.
Conflict of interest statement
Figures

References
-
- Food and Agriculture Organization of the United Nations website. Available: http://www.fao.org/docrep/012/al390e/al390e00.pdf. Accessed 2011 April 19.
-
- Barker DJ. Maternal nutrition and cardiovascular disease. Nutr Health. 1993;9:99–106. - PubMed
-
- Barker DJ. Mothers, babies, and disease in later life. London: BMJ Publishing Group; 1994. 232
-
- Barker DJ. Fetal nutrition and cardiovascular disease in later life. British Med Bull. 1997;53:96–108. - PubMed
-
- Barker DJ. Fetal programming of coronary heart disease. Endocrinology & Metabolism. 2002;13:364–368. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
