

Dysregulation of the unfolded protein response in db/db mice with diet-induced steatohepatitis
- PMID: 21748768
- PMCID: PMC3205284
- DOI: 10.1002/hep.24553
Dysregulation of the unfolded protein response in db/db mice with diet-induced steatohepatitis
Abstract
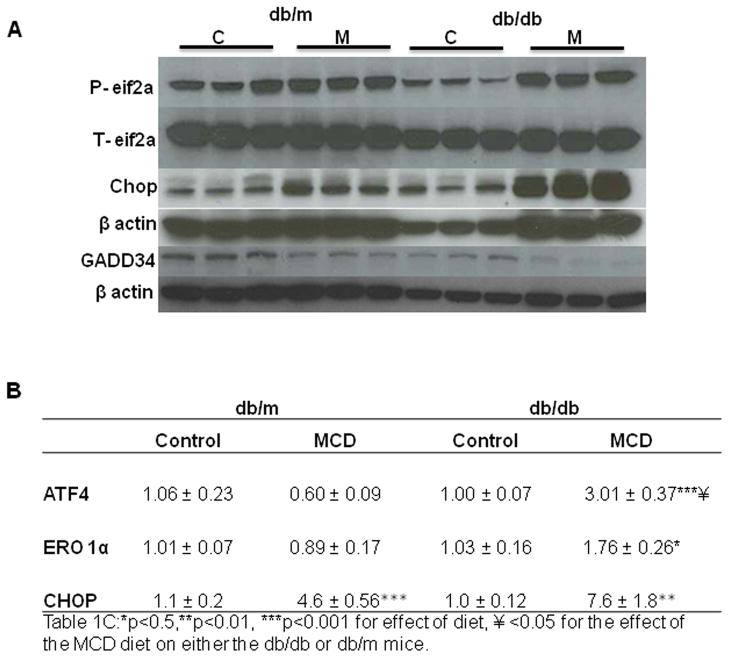
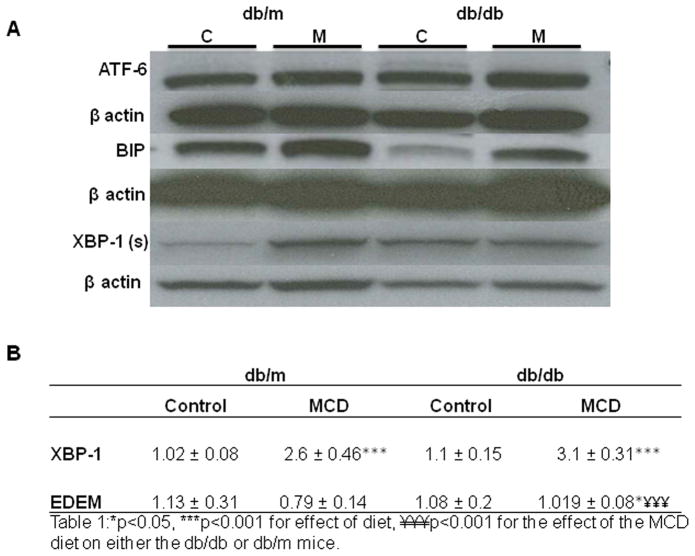
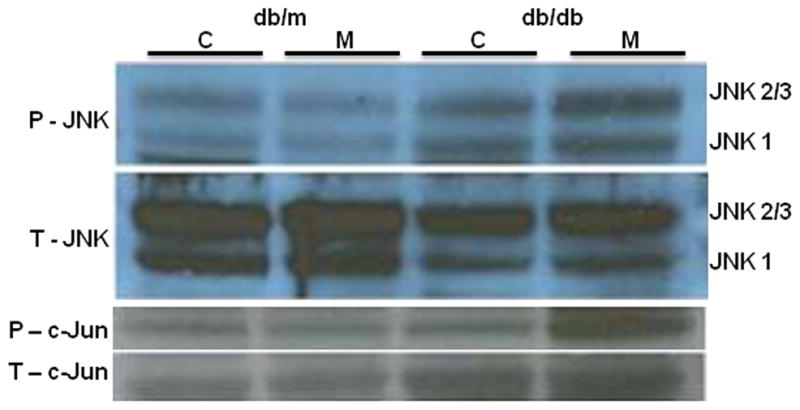
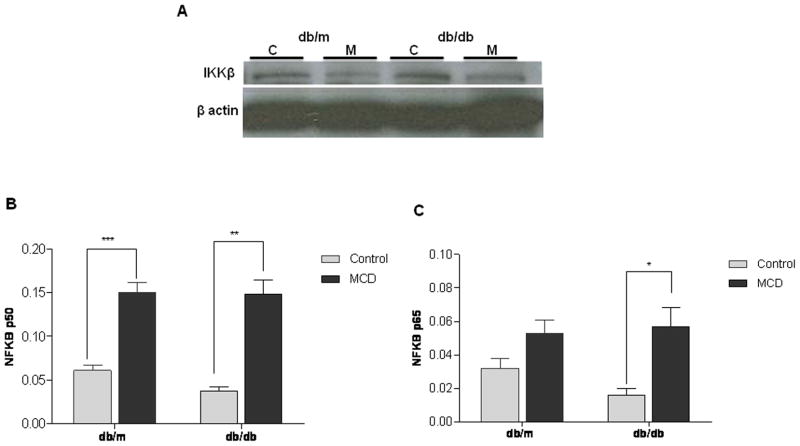
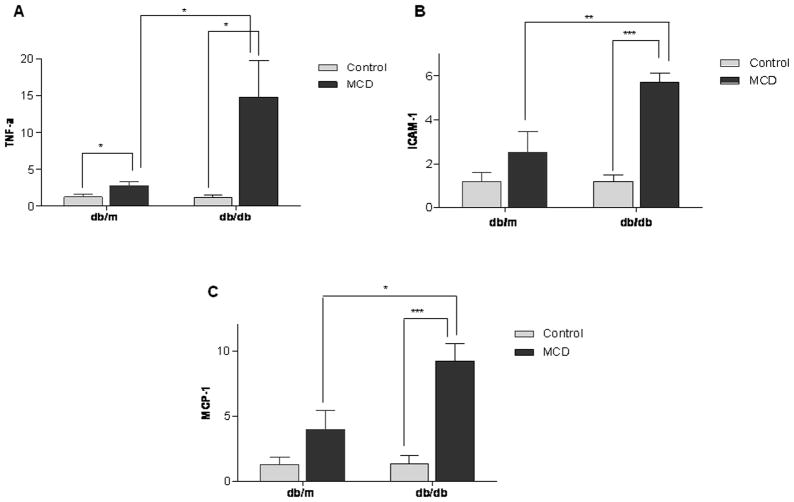
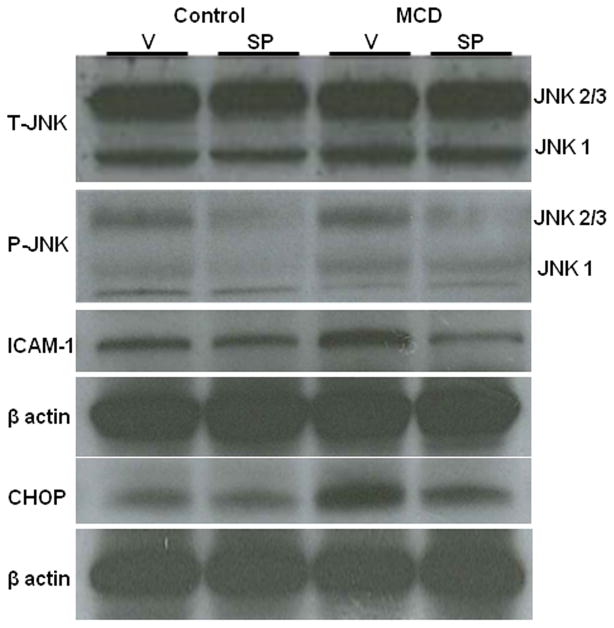
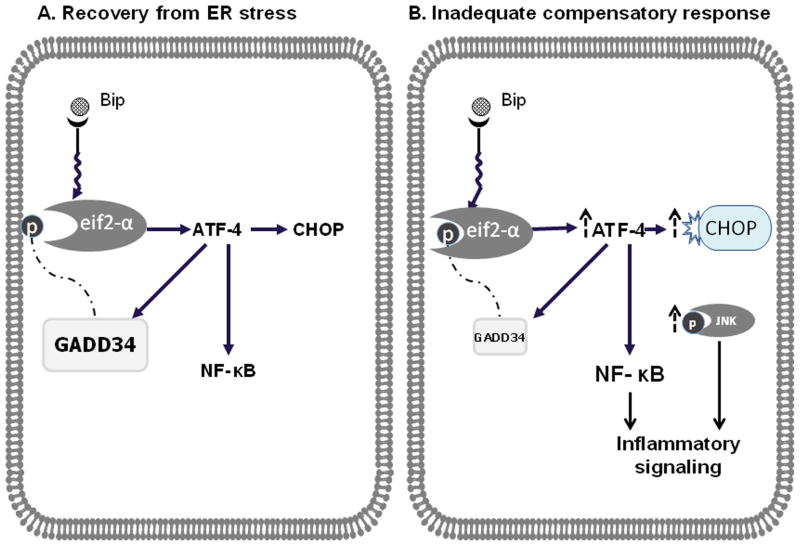
In humans with nonalcoholic fatty liver, diabetes is associated with more advanced disease. We have previously shown that diabetic db/db mice are highly susceptible to methionine choline-deficient diet (MCD)-induced hepatic injury. Because activation of the unfolded protein response (UPR) is an important adaptive cellular mechanism in diabetes, obesity, and fatty liver, we hypothesized that dysregulation of the UPR may partially explain how diabetes could promote liver injury. Db/db and db/m mice were fed the MCD or control diet for 4 weeks to characterize differences in UPR activation and downstream injury. Wildtype mice (C57BLKS/J) fed the MCD or control diet were treated with SP600125; a c-Jun N-terminal kinase (JNK) inhibitor and its effect on liver injury and UPR activation was measured. The MCD diet resulted in global up-regulation of the UPR in both diabetic db/db and nondiabetic db/m mice. db/db mice had an inadequate activation of recovery pathways (GADD34, XBP-1(s)) and accentuated activation of injury pathways related to persistent eif2-α phosphorylation (activating transcription factor 4 [ATF-4], C/EBP homologous transcription factor [CHOP], oxireductase endoplasmic reticulum oxidoreductin-1 [ERO-1α], JNK, nuclear factor kappaB [NF-κB]) compared to db/m mice. This led to increased expression of inflammatory mediators such as tumor necrosis factor alpha (TNF-α), ICAM-1, and MCP-1 compared to db/m mice. Interestingly, whereas pharmacologic JNK inhibition did not prevent the development of MCD diet-induced steatohepatitis, it did attenuate UPR and downstream inflammatory signaling.
Conclusion: MCD-fed db/db mice develop a more proinflammatory milieu than db/m mice associated with an impaired ability to dephosphorylate eif2-α through GADD34, impairing cellular recovery. These data may enhance our understanding of why diabetics with nonalcoholic steatohepatitis are prone to develop more severe liver injury than nondiabetic patients.
Copyright © 2011 American Association for the Study of Liver Diseases.
Figures
References
-
- Czaja MJ. Liver injury in the setting of steatosis: crosstalk between adipokine and cytokine. Hepatology. 2004;40:19–22. - PubMed
-
- Samuel VT, Liu ZX, Qu X, Elder BD, Bilz S, Befroy D, et al. Mechanism of hepatic insulin resistance in non-alcoholic fatty liver disease. J Biol Chem. 2004;279:32345–32353. - PubMed
-
- Rinella ME, Green RM. The methionine-choline deficient dietary model of steatohepatitis does not exhibit insulin resistance. J Hepatol. 2004;40:47–51. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous