

Osteomyelosclerosis, anemia and extramedullary hematopoiesis in mice lacking the transcription factor NFATc2
- PMID: 21750088
- PMCID: PMC3208674
- DOI: 10.3324/haematol.2011.042515
Osteomyelosclerosis, anemia and extramedullary hematopoiesis in mice lacking the transcription factor NFATc2
Abstract
Background: Nuclear factors of activated T cells (NFAT) are transcription factors that are central to cytokine production in activated T cells and regulate the development and differentiation of various tissues. NFATc2 is expressed in hematopoietic stem cells and regulated during myeloid commitment in a lineage-specific manner. The biological role of NFATc2 in hematopoiesis is, however, unclear.
Design and methods: In the present study, we analyzed steady-state hematopoiesis in young (<3 months) and old (>12 months) mice lacking NFATc2. Complete blood counts were performed in the peripheral blood, bone marrow and spleen. Using cytological and histological analyses, the blood cell differential was determined. Colony-formation assays were used to determine the differentiation potential of hematopoietic cells. Bone cell cultures were derived from the bone marrow, and bone remodeling markers were determined in the serum.
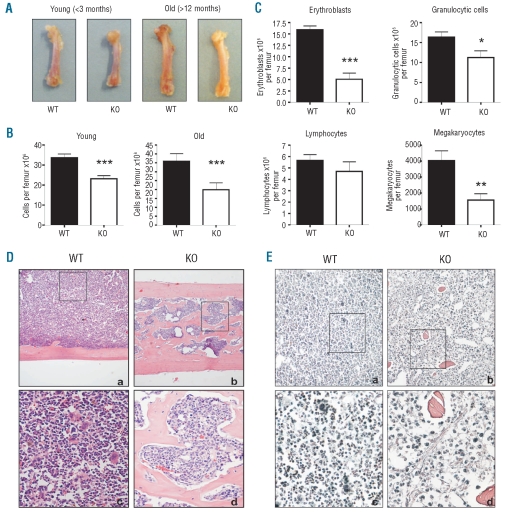
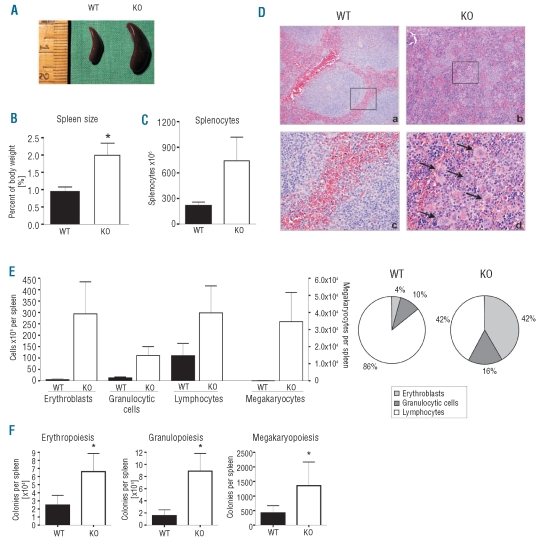
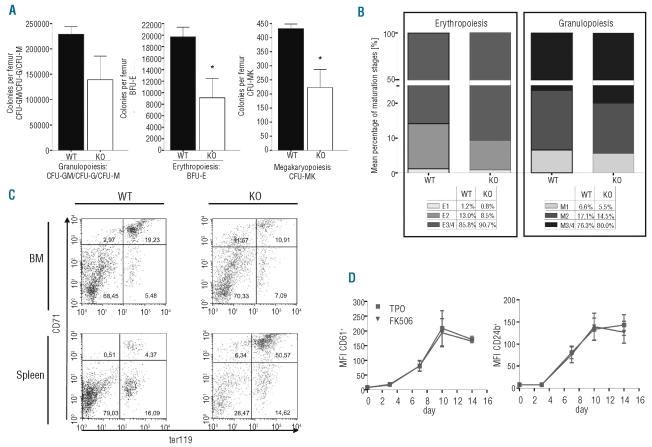
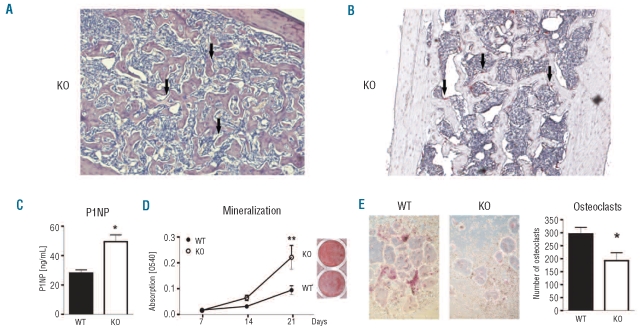
Results: NFATc2(-/-) mice older than 12 months were anemic and thrombocytopenic. The bone marrows of these mice showed a markedly reduced number of hematopoietic cells, of which megakaryocytic and erythroid lineages were most affected. While the number of hematopoietic progenitor cells in NFATc2-deficent bone marrow was reduced, the myeloid differentiation potential of these cells remained intact. Aged NFATc2(-/-) mice showed ossification of their bone marrow space and developed extramedullary hematopoiesis in the spleen. Ex vivo differentiation assays revealed an intrinsic defect of NFATc2-deficient stromal cells, in which NFATc2(-/-) osteoblasts differentiated more efficiently than wild-type cells, whereas osteoclast differentiation was impaired.
Conclusions: Our data suggest that NFATc2 may play a role in the maintenance of steady-state hematopoiesis and bone remodeling in adult organisms.
Figures
References
-
- Orkin SH. Diversification of haematopoietic stem cells to specific lineages. Nat Rev Genet. 2000;1(1):57–64. - PubMed
-
- Kiel MJ, Morrison SJ. Uncertainty in the niches that maintain haematopoietic stem cells. Nat Rev Immunol. 2008;8(4):290–301. - PubMed
-
- Calvi LM, Adams GB, Weibrecht KW, Weber JM, Olson DP, Knight MC, et al. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature. 2003;425(6960):841–6. - PubMed
-
- Majka M, Janowska-Wieczorek A, Ratajczak J, Ehrenman K, Pietrzkowski Z, Kowalska MA, et al. Numerous growth factors, cytokines, and chemokines are secreted by human CD34(+) cells, myeloblasts, erythroblasts, and megakaryoblasts and regulate normal hematopoiesis in an autocrine/paracrine manner. Blood. 2001;97(10):3075–85. - PubMed
-
- Zhang J, Niu C, Ye L, Huang H, He X, Tong WG, et al. Identification of the haematopoietic stem cell niche and control of the niche size. Nature. 2003;425(6960):836–41. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
