

Unique cell adhesion and invasion properties of Yersinia enterocolitica O:3, the most frequent cause of human Yersiniosis
- PMID: 21750675
- PMCID: PMC3131269
- DOI: 10.1371/journal.ppat.1002117
Unique cell adhesion and invasion properties of Yersinia enterocolitica O:3, the most frequent cause of human Yersiniosis
Abstract
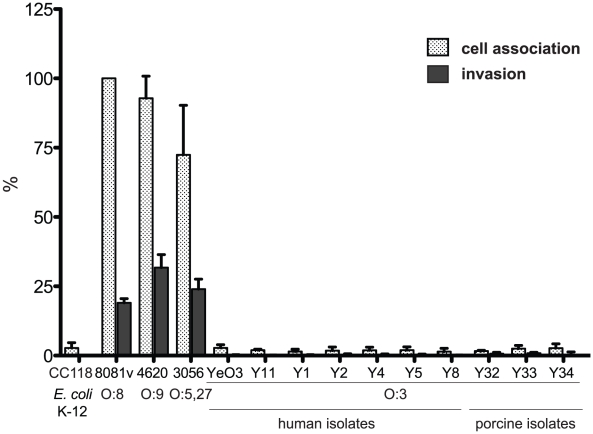
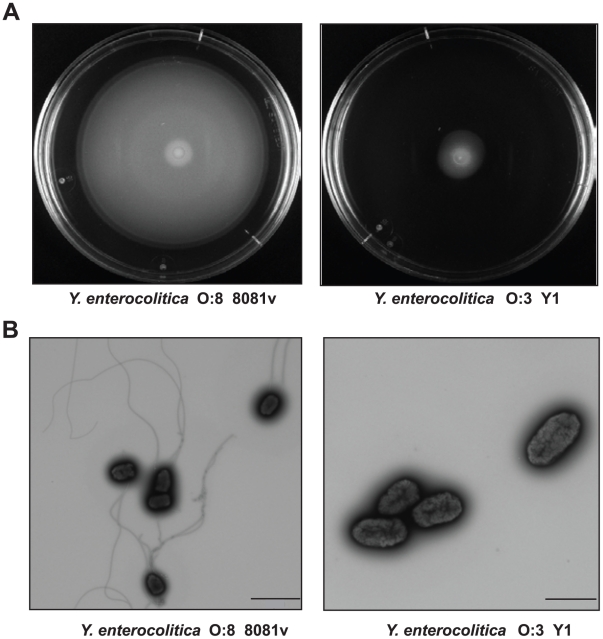
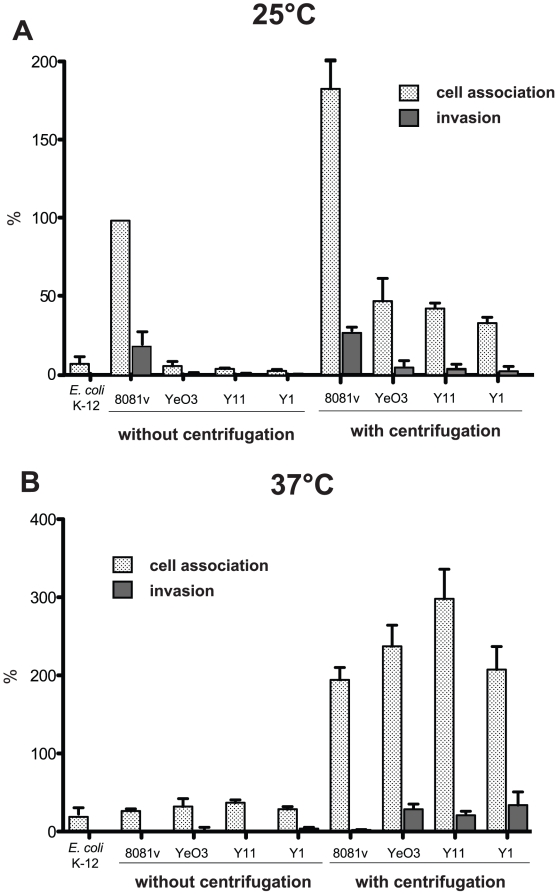
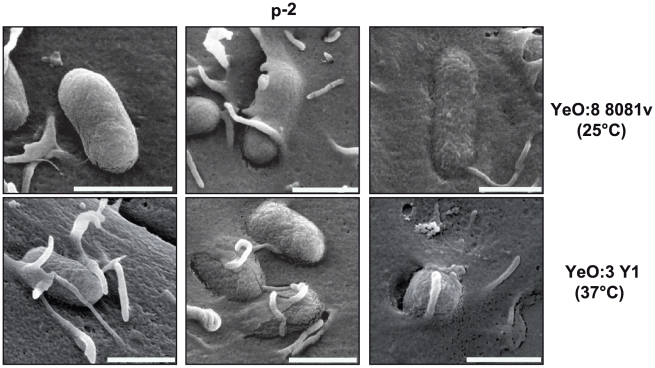
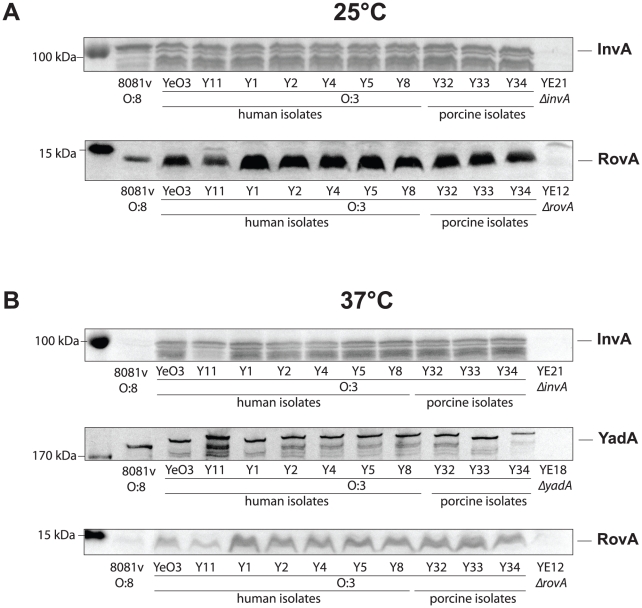
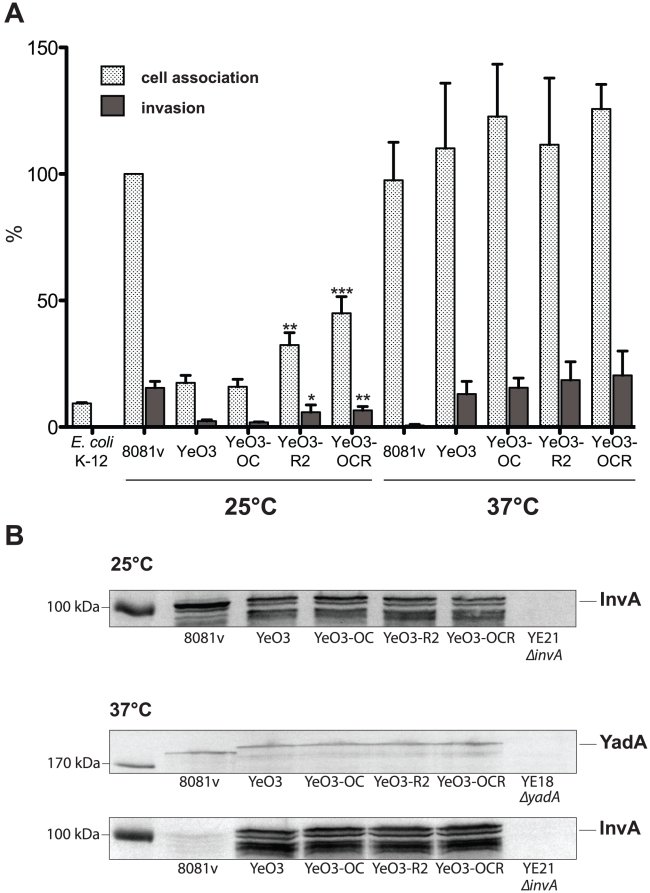
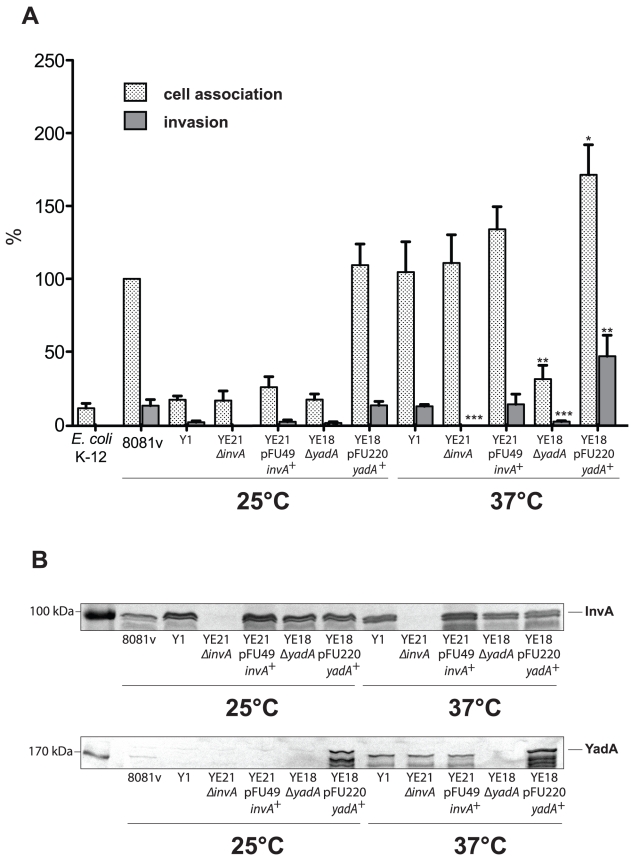
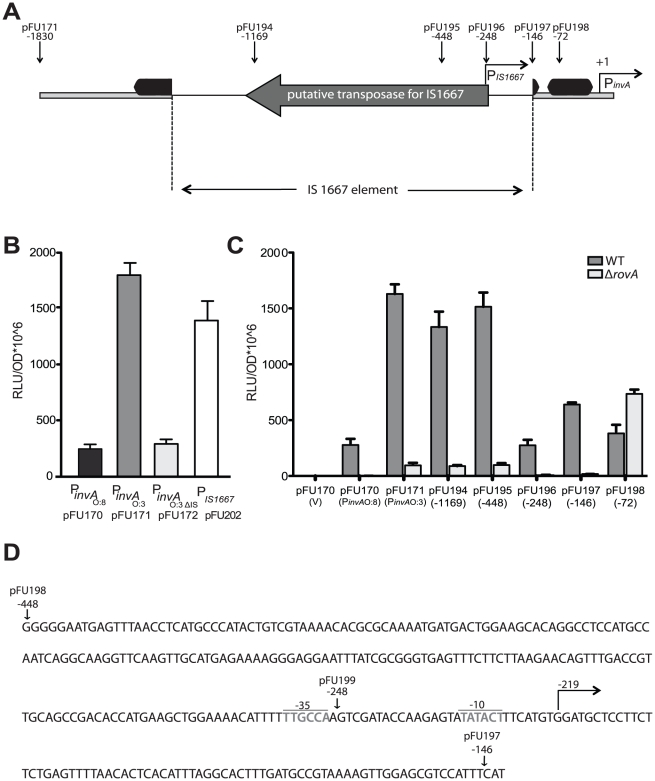
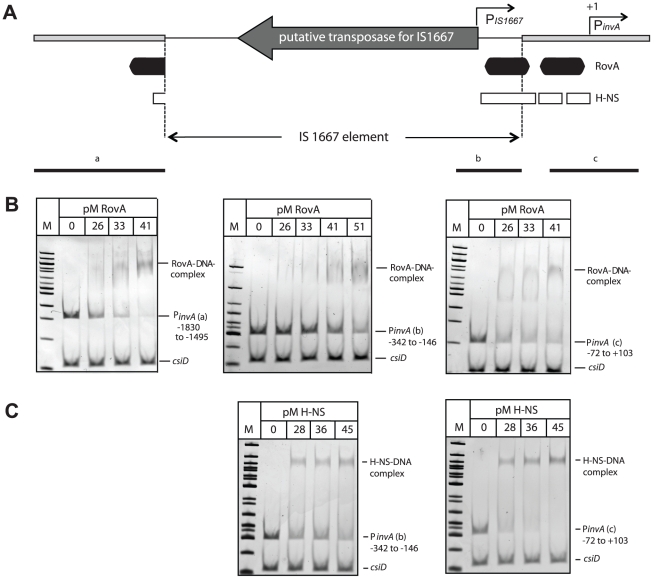
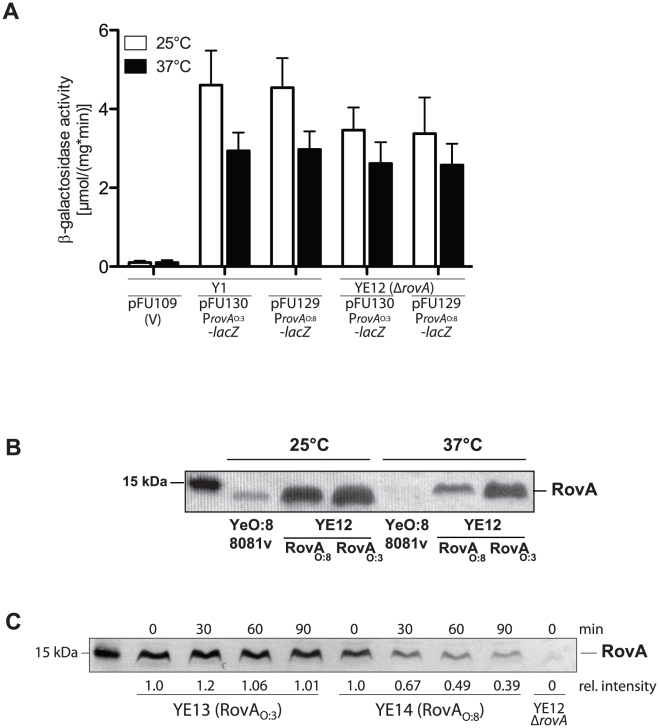
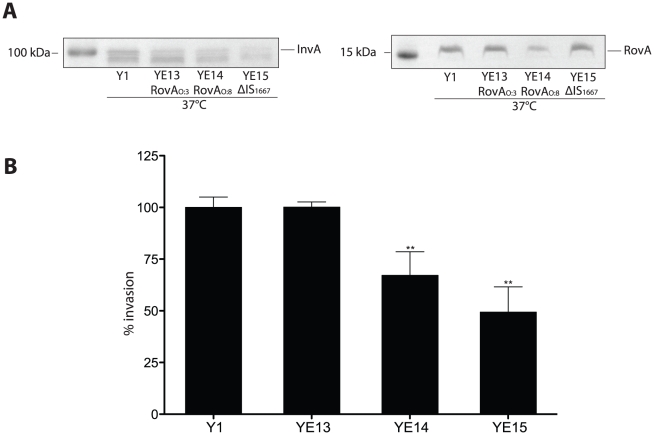
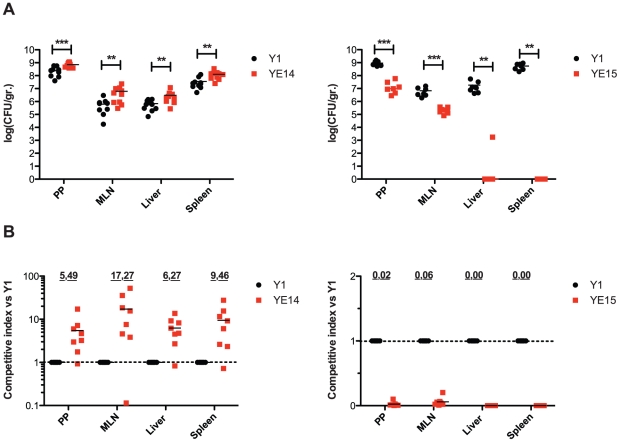
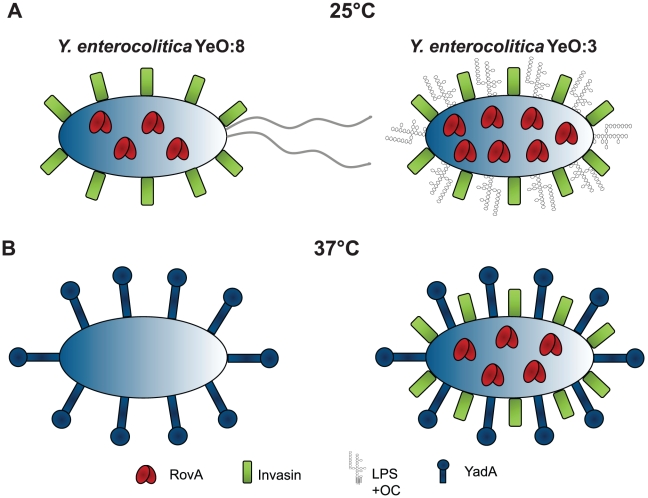
Many enteric pathogens are equipped with multiple cell adhesion factors which are important for host tissue colonization and virulence. Y. enterocolitica, a common food-borne pathogen with invasive properties, uses the surface proteins invasin and YadA for host cell binding and entry. In this study, we demonstrate unique cell adhesion and invasion properties of Y. enterocolitica serotype O:3 strains, the most frequent cause of human yersiniosis, and show that these differences are mainly attributable to variations affecting the function and expression of invasin in response to temperature. In contrast to other enteric Yersinia strains, invasin production in O:3 strains is constitutive and largely enhanced compared to other Y. enterocolitica serotypes, in which invA expression is temperature-regulated and significantly reduced at 37°C. Increase of invasin levels is caused by (i) an IS1667 insertion into the invA promoter region, which includes an additional promoter and RovA and H-NS binding sites, and (ii) a P98S substitution in the invA activator protein RovA rendering the regulator less susceptible to proteolysis. Both variations were shown to influence bacterial colonization in a murine infection model. Furthermore, we found that co-expression of YadA and down-regulation of the O-antigen at 37°C is required to allow efficient internalization by the InvA protein. We conclude that even small variations in the expression of virulence factors can provoke a major difference in the virulence properties of closely related pathogens which may confer better survival or a higher pathogenic potential in a certain host or host environment.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Hanski C, Naumann M, Hahn H, Riecken EO. Determinants of invasion and survival of Yersinia enterocolitica in intestinal tissue. An in vivo study. Med Microbiol Immunol. 1989;178:289–296. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
