

Both the C-terminal polylysine region and the farnesylation of K-RasB are important for its specific interaction with calmodulin
- PMID: 21750741
- PMCID: PMC3130059
- DOI: 10.1371/journal.pone.0021929
Both the C-terminal polylysine region and the farnesylation of K-RasB are important for its specific interaction with calmodulin
Abstract
Background: Ras protein, as one of intracellular signal switches, plays various roles in several cell activities such as differentiation and proliferation. There is considerable evidence showing that calmodulin (CaM) binds to K-RasB and dissociates K-RasB from membrane and that the inactivation of CaM is able to induce K-RasB activation. However, the mechanism for the interaction of CaM with K-RasB is not well understood.
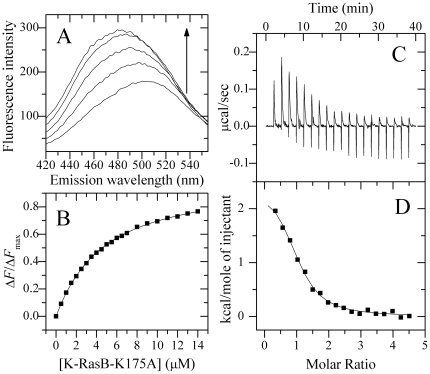
Methodology/principal findings: Here, by applying fluorescence spectroscopy and isothermal titration calorimetry, we have obtained thermodynamic parameters for the interaction between these two proteins and identified the important elements of K-RasB for its interaction with Ca(2+)/CaM. One K-RasB molecule interacts with one CaM molecule in a GTP dependent manner with moderate, micromolar affinity at physiological pH and physiologic ionic strength. Mutation in the polybasic domain of K-Ras decreases the binding affinity. By using a chimera in which the C-terminal polylysine region of K-RasB has been replaced with that of H-Ras and vice versa, we find that at physiological pH, H-Ras-(KKKKKK) and Ca(2+)/CaM formed a 1:1 complex with an equilibrium association constant around 10(5) M(-1), whereas no binding reaction of K-RasB-(DESGPC) with Ca(2+)/CaM is detected. Furthermore, the interaction of K-RasB with Ca(2+)/CaM is found to be enhanced by the farnesylation of K-RasB.
Conclusions/significance: We demonstrate that the polylysine region of K-RasB not only contributes importantly to the interaction of K-RasB with Ca(2+)/CaM, but also defines its isoform specific interaction with Ca(2+)/CaM. The farnesylation of K-RasB is also important for its specific interaction with Ca(2+)/CaM. Information obtained here can enhance our understanding of how CaM interacts with K-RasB in physiological environments.
Conflict of interest statement
Figures





References
-
- Barbacid M. ras genes. Annu Rev Biochem. 1987;56:779–827. - PubMed
-
- Bos JL. ras oncogenes in human cancer: a review. Cancer Res. 1989;49:4682–4689. - PubMed
-
- Downward J. Targeting Ras signalling pathways in cancer therapy. Nat Rev Cancer. 2003;3:11–22. - PubMed
-
- Hancock JF. Ras proteins:different signals from different locations. Nat Rev Mol Cell Biol. 2003;4:373–384. - PubMed
-
- Klee CB, Crouch TH, Richman PG. Calmodulin. Annu Rev Biochem. 1980;49:489–515. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
