

Zoledronic acid inhibits macrophage SOCS3 expression and enhances cytokine production
- PMID: 21751240
- PMCID: PMC3196771
- DOI: 10.1002/jcb.23267
Zoledronic acid inhibits macrophage SOCS3 expression and enhances cytokine production
Abstract
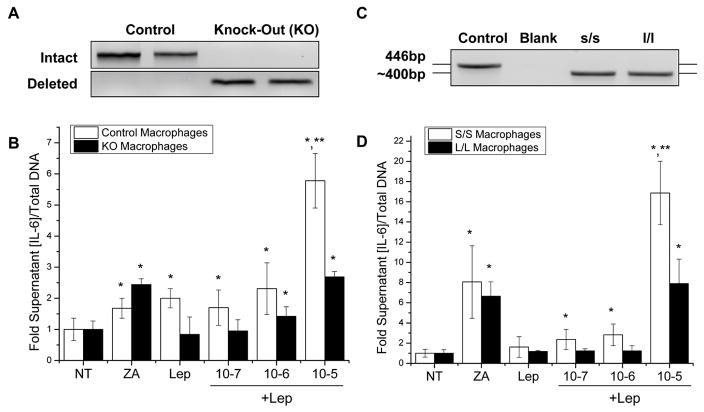
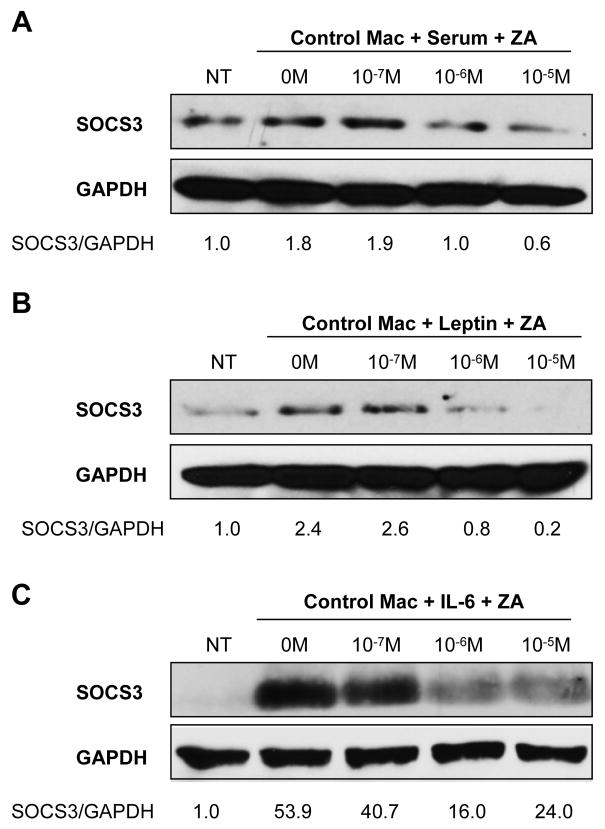
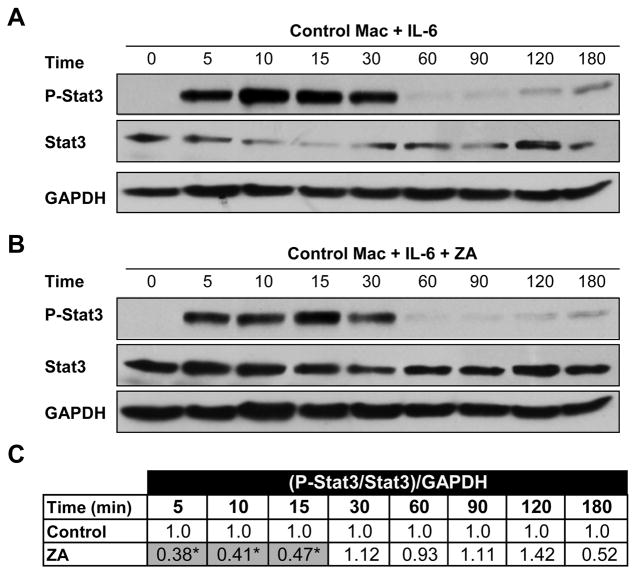
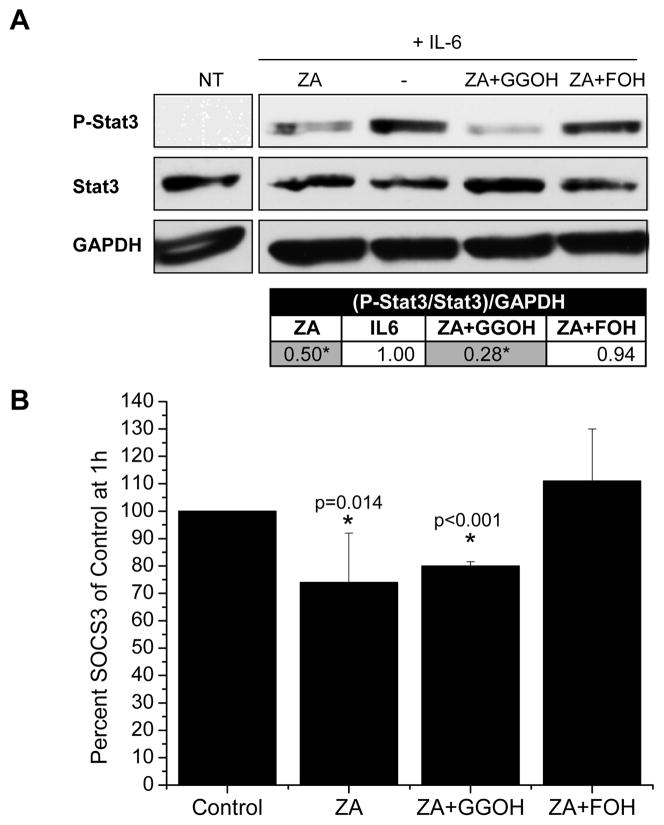
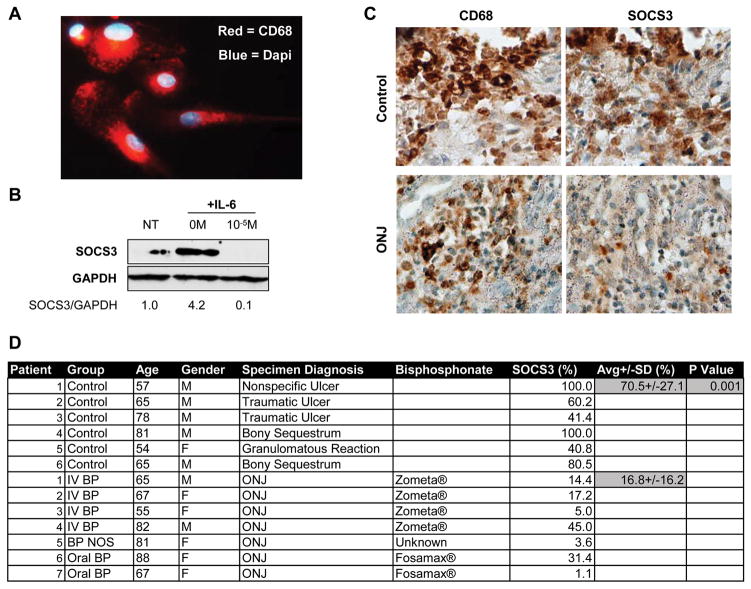
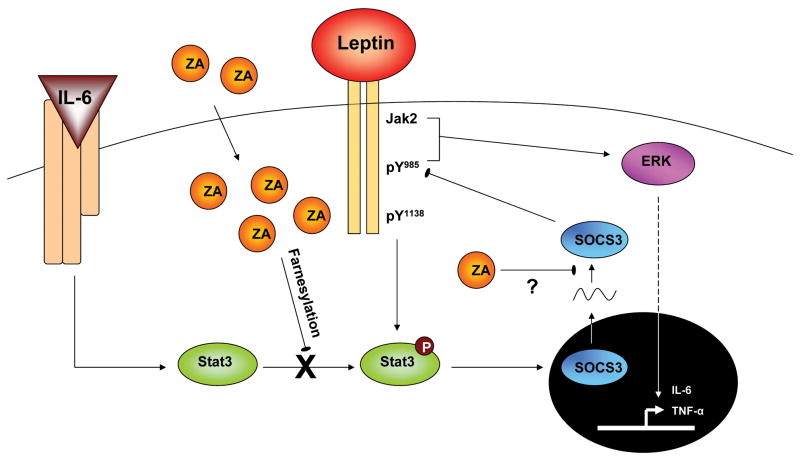
Suppressor of cytokine signaling-3 (SOCS3) has multiple functions including inhibition of Janus kinase (Jak) activity, regulation of protein degradation, and suppression of cytokine signaling. SOCS3 modulates macrophage response to cytokines such as IL-6 and leptin that are systemically induced in obesity. Obesity is a suspected risk factor for SOCS3-related pathology such as rheumatoid arthritis and Crohn's disease as well as zoledronic acid (ZA)-induced osteonecrosis of the jaw (ONJ). Thus, understanding the ability of bisphosphonates to modulate SOCS3 is necessary to qualify their contribution to these disorders. ONJ occurs in up to 10% of patients using intravenous bisphosphonates and has an unknown pathogenesis that may be linked to decreased bone turnover, altered vascularity, bacterial invasion, and compromised wound healing. Given the increased risk of ONJ with obesity and importance of macrophages in wound healing, we hypothesized that amino-bisphosphonates could contribute to the pathogenesis of ONJ by regulating macrophage responses to cytokines such as leptin and IL-6. We report that ZA is a novel inhibitor of SOCS3 in primary macrophages and human ONJ biopsy specimens. Inhibition of SOCS3 by ZA resulted in significant increases in IL-6 production. SOCS3 transcription is regulated by nuclear accumulation of phosphorylated-Stat3 (P-Stat3). We found that ZA decreased phosphorylation of Stat3 in a mevalonate-pathway dependent manner. However, restoration of P-Stat3 was not sufficient to correct SOCS3 inhibition. We propose that disruption of macrophage SOCS3 expression by amino-bisphosphonates such as ZA may be a novel contributor to inflammatory phenotypes in obesity and the pathogenesis of ONJ.
Copyright © 2011 Wiley Periodicals, Inc.
Figures

References
-
- Banks AS, Davis SM, Bates SH, Myers MG., Jr Activation of downstream signals by the long form of the leptin receptor. J Biol Chem. 2000;275:14563–72. - PubMed
-
- Bates SH, Stearns WH, Dundon TA, Schubert M, Tso AW, Wang Y, Banks AS, Lavery HJ, Haq AK, Maratos-Flier E, Neel BG, Schwartz MW, Myers MG., Jr STAT3 signalling is required for leptin regulation of energy balance but not reproduction. Nature. 2003;421:856–9. - PubMed
-
- Cartsos VM, Zhu S, Zavras AI. Bisphosphonate use and the risk of adverse jaw outcomes: a medical claims study of 714,217 people. J Am Dent Assoc. 2008;139:23–30. - PubMed
-
- Chang MK, Raggatt LJ, Alexander KA, Kuliwaba JS, Fazzalari NL, Schroder K, Maylin ER, Ripoll VM, Hume DA, Pettit AR. Osteal tissue macrophages are intercalated throughout human and mouse bone lining tissues and regulate osteoblast function in vitro and in vivo. J Immunol. 2008;181:1232–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
