

Cell-type-specific type I interferon antagonism influences organ tropism of murine coronavirus
- PMID: 21752905
- PMCID: PMC3196400
- DOI: 10.1128/JVI.05075-11
Cell-type-specific type I interferon antagonism influences organ tropism of murine coronavirus
Abstract
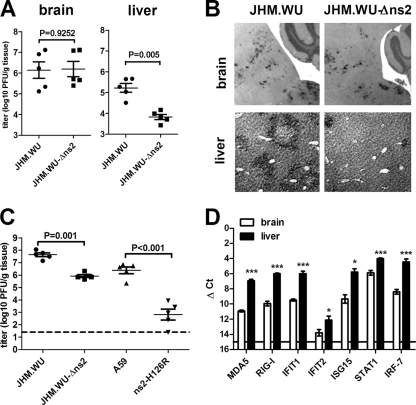
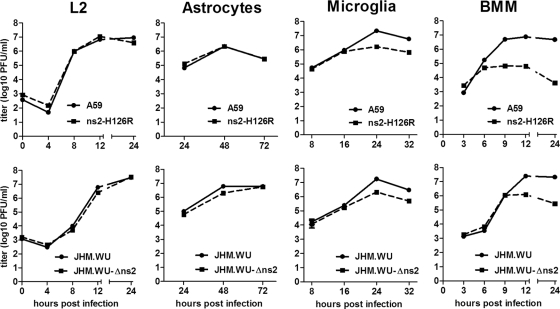
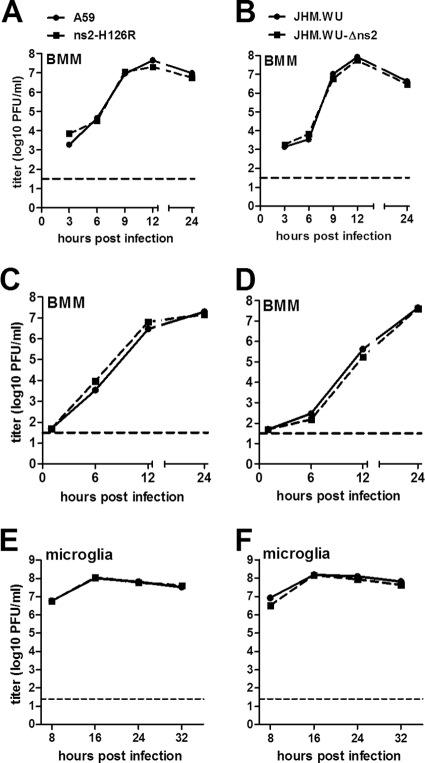
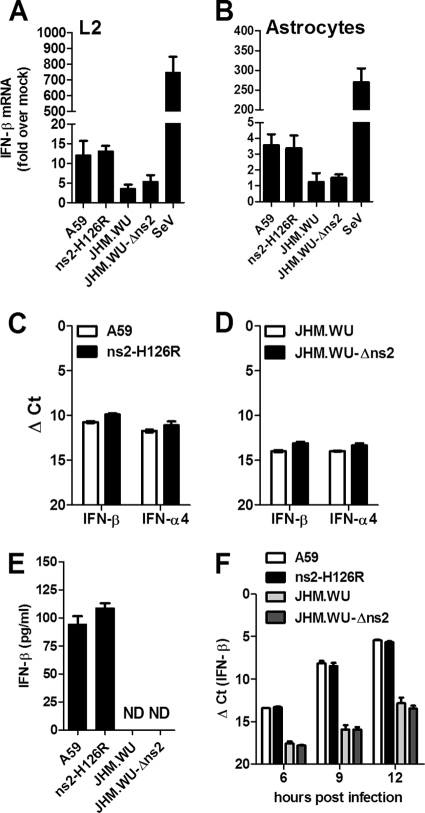
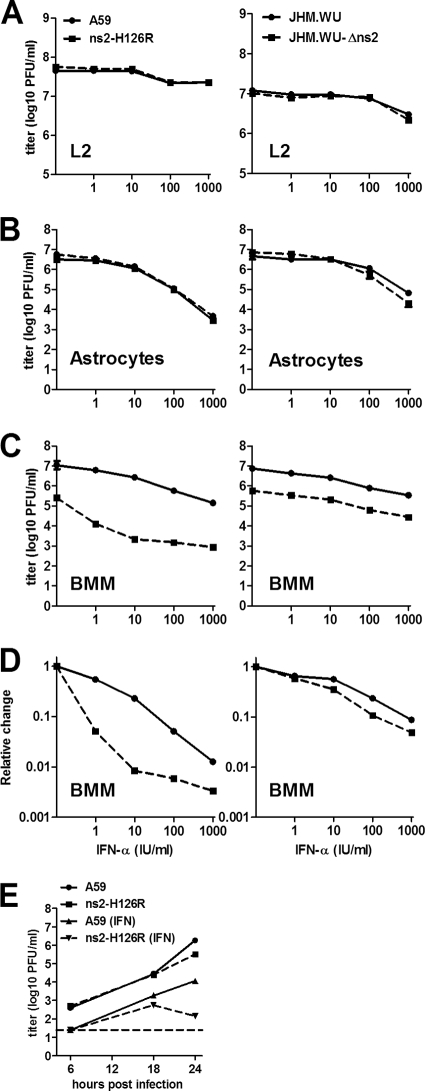
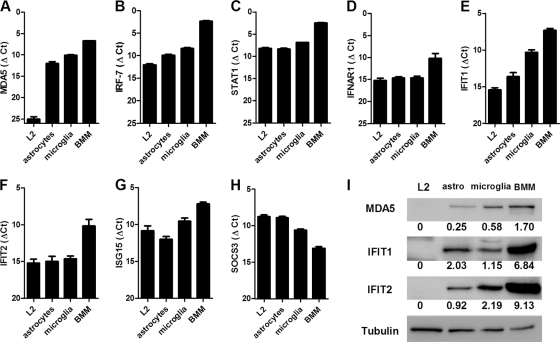
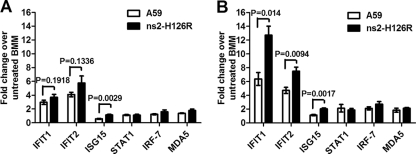
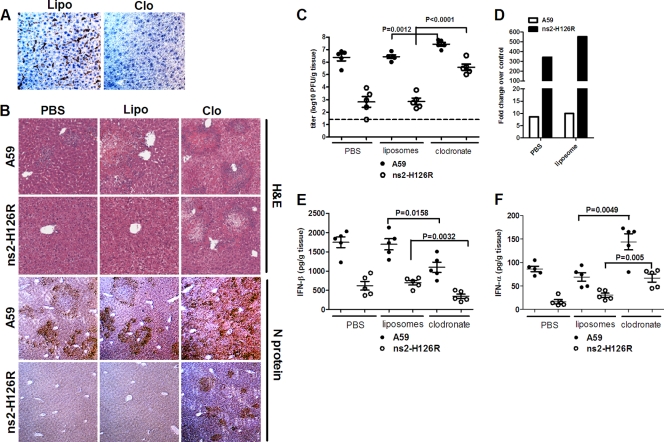
Previous studies have demonstrated that mouse hepatitis virus (MHV) hepatotropism is determined largely by postentry events rather than by availability of the viral receptor. In addition, mutation of MHV nonstructural protein 2 (ns2) abrogates the ability of the virus to replicate in the liver and induce hepatitis but does not affect replication in the central nervous system (CNS). Here we show that replication of ns2 mutant viruses is attenuated in bone marrow-derived macrophages (BMM) generated from wild-type (wt) mice but not in L2 fibroblasts, primary astrocytes, or BMM generated from type I interferon receptor-deficient (IFNAR(-/-)) mice. In addition, ns2 mutants are more sensitive than wt virus to pretreatment of BMM, but not L2 fibroblasts or primary astrocytes, with alpha/beta interferon (IFN-α/β). The ns2 mutants induced similar levels of IFN-α/β in wt and IFNAR(-/-) BMM, indicating that ns2 expression has no effect on the induction of IFN but rather that it antagonizes a later step in IFN signaling. Consistent with these in vitro data, the virulence of ns2 mutants increased to near that of wt virus after depletion of macrophages in vivo. These data imply that the ability of MHV to replicate in macrophages is a prerequisite for replication in the liver and induction of hepatitis but not for replication or disease in the CNS, underscoring the importance of IFN signaling in macrophages in vivo for protection of the host from hepatitis. Our results further support the notion that viral tissue tropism is determined in part by postentry events, including the early type I interferon response.
Figures
References
-
- Caamano J., Alexander J., Craig L., Bravo R., Hunter C. A. 1999. The NF-kappa B family member RelB is required for innate and adaptive immunity to Toxoplasma gondii. J. Immunol. 163:4453–4461 - PubMed
-
- Cervantes-Barragan L., et al. 2009. Type I IFN-mediated protection of macrophages and dendritic cells secures control of murine coronavirus infection. J. Immunol. 182:1099–1106 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
