

A dual strategy for the suppression of host antiviral silencing: two distinct suppressors for viral replication and viral movement encoded by potato virus M
- PMID: 21752911
- PMCID: PMC3196401
- DOI: 10.1128/JVI.05273-11
A dual strategy for the suppression of host antiviral silencing: two distinct suppressors for viral replication and viral movement encoded by potato virus M
Abstract
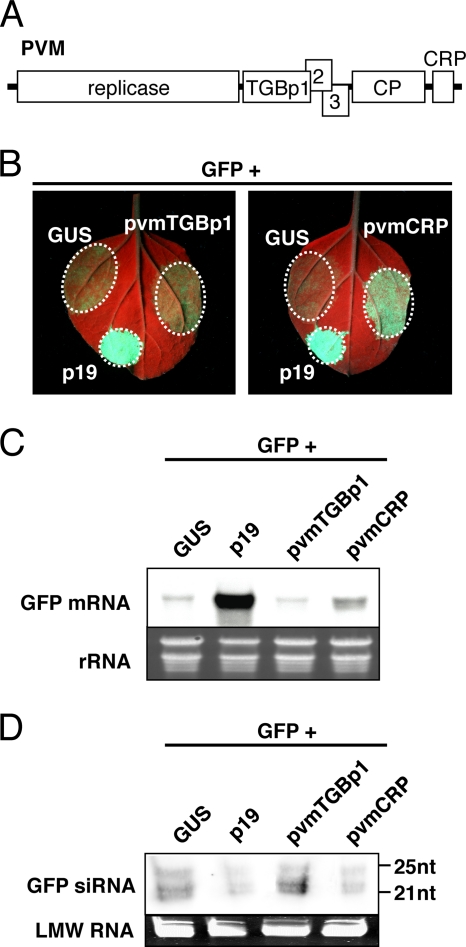
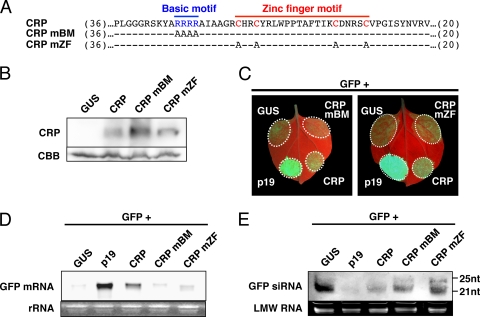
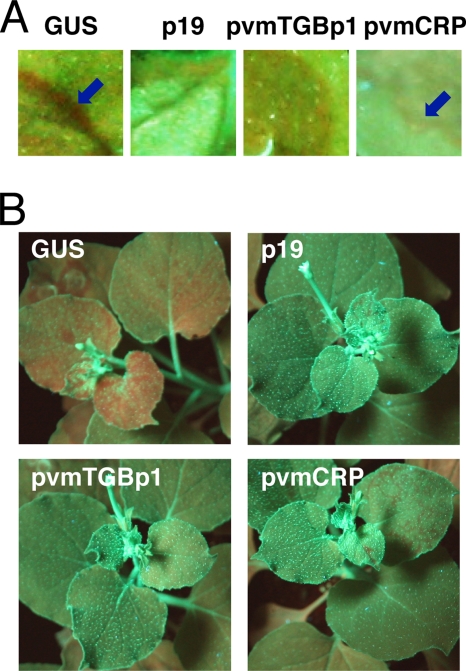
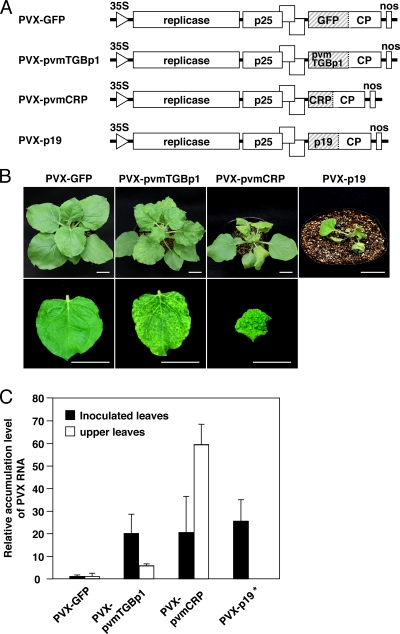
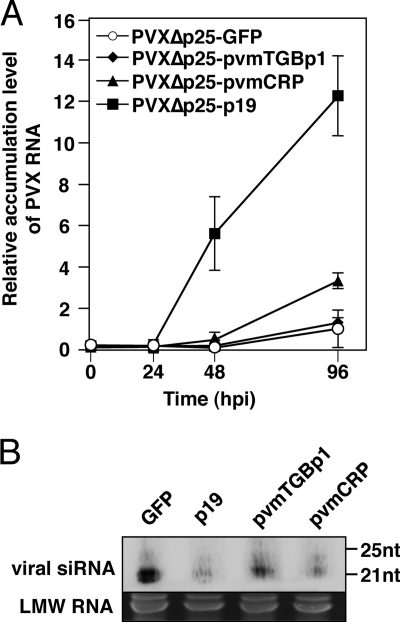
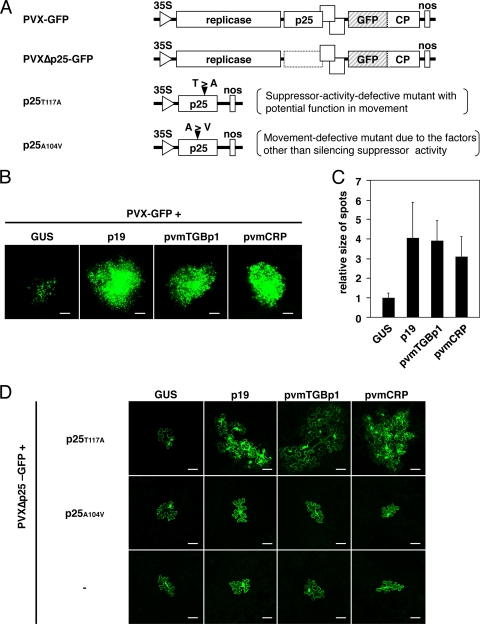
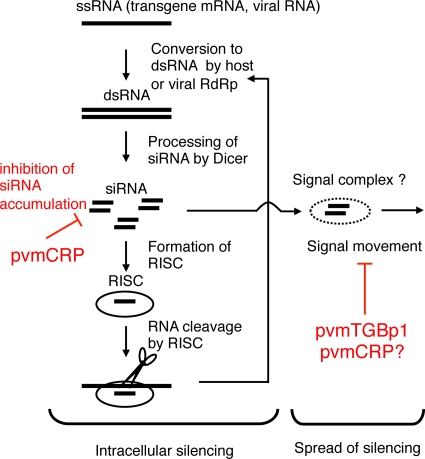
Viruses encode RNA silencing suppressors to counteract host antiviral silencing. In this study, we analyzed the suppressors encoded by potato virus M (PVM), a member of the genus Carlavirus. In the conventional green fluorescent protein transient coexpression assay, the cysteine-rich protein (CRP) of PVM inhibited both local and systemic silencing, whereas the triple gene block protein 1 (TGBp1) showed suppressor activity only on systemic silencing. Furthermore, to elucidate the roles of these two suppressors during an active viral infection, we performed PVX vector-based assays and viral movement complementation assays. CRP increased the accumulation of viral RNA at the single-cell level and also enhanced viral cell-to-cell movement by inhibiting RNA silencing. However, TGBp1 facilitated viral movement but did not affect viral accumulation in protoplasts. These data suggest that CRP inhibits RNA silencing primarily at the viral replication step, whereas TGBp1 is a suppressor that acts at the viral movement step. Thus, our findings demonstrate a sophisticated viral infection strategy that suppresses host antiviral silencing at two different steps via two mechanistically distinct suppressors. This study is also the first report of the RNA silencing suppressor in the genus Carlavirus.
Figures
References
-
- Baulcombe D. 2004. RNA silencing in plants. Nature 431:356–363 - PubMed
-
- Baumberger N., Tsai C. H., Lie M., Havecker E., Baulcombe D. C. 2007. The polerovirus silencing suppressor P0 targets Argonaute proteins for degradation. Curr. Biol. 17:1609–1614 - PubMed
-
- Bayne E. H., Rakitina D. V., Morozov S. Y., Baulcombe D. C. 2005. Cell-to-cell movement of potato potexvirus X is dependent on suppression of RNA silencing. Plant J. 44:471–482 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
